 2003, Vol. 39
2003, Vol. 39文章信息
- 董丽芬, 邵崇斌, 张宗勤.
- Dong Lifen, Shao Chongbin, Zhang Zongqin.
- 白皮松种胚的休眠萌发特性研究
- CHARACTERISTICS OF EMBRYO DORMANCY AND GERMINATION OF PINUS BUNGEANA
- 林业科学, 2003, 39(6): 47-54.
- Scientia Silvae Sinicae, 2003, 39(6): 47-54.
-
文章历史
- 收稿日期:2002-04-27
-
作者相关文章


白皮松(Pinus bungeana)具有很高的经济、生态和观赏价值,但其种子具深休眠特性,目前对其休眠机理还缺乏深刻认识,在快速催芽方法的研究上存在盲目性。在松属休眠机理方面,有关激素、光敏素、营养物代谢、酶活性、种皮效应、低温层积等因素与种子萌发的关系研究较多(GOO et al.,1952;Stone,1957a;1957b;Asakawa,1961;Kao,1972;Barnett, 1972;王文章等,1979;1986;张良诚等, 1981;谭志一等,1983;傅家瑞, 1985;Hellum et al., 1988;Lammer et al.,1989;程淑婉等,1992;Gurt et al., 1998;Shen et al., 1999;Jordy et al., 2000)。种胚休眠萌发特性的研究方面(Jackson et al., 1965;Le, 1973a;1973b;1977;黄玉国, 1986;许绍惠等,1988;Dawidowica, 1989;Sambeek et al., 1989;Arrillaga et al., 1992;Siti et al.,2000;Chapman,2000;Widrlechner, 2000),松属种胚研究较少(Baron,1966;Kao, 1970)。就白皮松种子休眠而言,对营养物质代谢、种子休眠的地理变异、种皮效应、催芽方法等曾有研究(陶章安, 1956;于淑兰,1983;李晓洁等,1989;尚杰等,1992;Dong et al.,1993;赵焱等,1995;王小平等,1998;2000),但对胚的休眠与萌发特性研究甚少。研究白皮松种胚的休眠萌发特性,对认识白皮松种子休眠萌发的本质,避免种子快速催芽方法研究上的盲目性十分必要,同时对其它松属树种的研究也具借鉴意义。
1 材料与方法 1.1 试验材料白皮松种子来源于山西省乡宁县吕梁山台头林场,位于东经111°7′,北纬36°6′。采种母树生长在海拔1 200~1 600 m的半阴半阳坡,树龄20~40 a生。9月中旬至10月中旬种子完全成熟后采种。球果采用自然干燥、人工脱粒方法。试验用种是当年采收的新鲜种子,属同一种批。经TTC法测定(GB 2772-1999),生活力为99%。
1.2 试验方法 1.2.1 离体胚滤纸培养离体胚置于钟形培养皿的滤纸上,培养皿放在盛水的烧杯上,通过培养皿底盘孔的纱布条伸入烧杯的水中。胚的置床过程按无菌操作要求进行。整套离体胚滤纸培养装置放在恒温光照发芽箱中进行培养。每处理活胚培养总数大于或等于60个(表 1、表 2)。
|
|
|
|
根据预备试验,离体胚琼脂培养基成分设计如下。不加GA3的培养基:琼脂培养基+w%(w%示不同浓度的蔗糖,A:0%;B:0.1%;C:0.2%;D:0.4%;E:0.6%;F:0.8%;G:1.0%;H:1.5%;I:2.0%;J:2.5%;K:3.0%;L:4.0%;M:4.5%;N:5.0%;O:5.5%;P:6.0%;Q:8.0%;R:10.0%)。加GA3的培养基:琼脂培养基+GA3 (0.1 mg·L-1)+W'%(W'%示不同浓度的蔗糖,C':0.2%;D':0.4%;E':0.6%;F':0.8%;G':1.0%;I':2.0%;K':3.0%;L':4.0%;N':5.0%)。操作全过程按无菌操作要求进行。采用具刻度的5 mL试管,每管装入2 mL琼脂培养基。在培养基斜面中央,接种1个健康的离体胚,如图 1、2、3所示。把接过胚的试管置恒温光照发芽箱中培养。各处理培养活胚总数均不少于20个(表 1~3)。

|
图 1 非低温层积种子离体胚在蔗糖浓度为0.1%的琼脂培养基上的生长状态(下胚轴生长量较大,但胚根未萌发) Fig. 1 The growth of excised embryo of the seeds without cold stratification, incubated on the medium with sucrose at 0.1%(Hypocotyl elongated, and radicle did not germinate) |

|
图 2 非低温层积种子离体胚在蔗糖浓度为3.0%的琼脂培养基上的萌发状态 Fig. 2 The growth of excised embryos of the seeds without cold stratification, incubated on the medium with sucrose at 3.0% 右:萌发出胚根的离体胚;左:未萌发胚根的离体胚,下胚轴下端有一红棕色胚根套。Right: radicle germinated; Left: hypocotyl was crowned a red brown jacket, and radicle did not germinate. |

|
图 3 低温层积36 d种子离体胚在蔗糖浓度为3.0%的琼脂培养基上萌发长出的幼嫩胚根 Fig. 3 The growth of excised embryos of the seeds treated by cold stratification for 36 days, incubated on medium with sucrose at 3.0%(longer and tender roots) |
|
|
培养装置同离体胚滤纸培养。按无菌操作要求把不带内外种皮的带全胚乳胚接种在培养皿的滤纸上,整套培养装置放入恒温光照发芽箱中培养。带部分胚乳胚是削去了1/3或1/2或2/3的胚乳,培养方法同全胚乳胚培养法。各处理培养数量均为150个(表 4)。
|
|
方法1:种子无预处理。指种子不进行任何方法的处理。方法2:种子低温层积36 d。把粒径为0.5~1 mm的河沙,用自来水冲洗掉泥土及粉沙,置烘箱中130℃烘2 h,加自来水搅匀,使沙子含水量为其最大持水量的60%。所用种子为无预处理的干种子。按沙:种为3:1(体积比)的比例把湿沙和干种混合均匀。把种沙混合体装入口径为8 cm容器中,在混合体上覆3 cm厚的纯湿沙。容器口不加盖,置5℃冷藏箱中。
1.2.5 恒温光照发芽箱的培养条件所有离体胚的滤纸培养及琼脂培养、带全胚乳胚及带部分胚乳胚的滤纸培养均放在同一恒温光照发芽箱中。温度控制在25℃±1℃,相对湿度保持在90%以上。距日光灯40 cm处,光强为850 lx。培养物还可利用室内自然光照,夏季睛天中午光强约4 500 lx。
1.2.6 试验种子抽样方法用四分法(GB 2772-1999)按所需量从大量种子中随机抽取各项试验种子。
1.2.7 试验观察记录方法胚萌发观察记录标准:离体胚萌发的幼根是该胚长度一半以上,带胚乳胚萌发的幼根是该胚乳长度一半以上。生长量测定:离体胚及带1/3、1/2或2/3胚乳胚的下胚轴伸长后或胚根萌发后2~3 d内停止生长,均在生长停止后测定生长量。带全胚乳胚生长量在胚乳从子叶上脱掉时测定。测定方法是把带部分胚乳的胚和琼脂培养基上的离体胚从胚乳中或培养基上取出,脱掉胚乳的带全胚乳胚用自来水小心冲冼干净,吸水纸吸干表面水分,按不同实验处理摆放在白纸上。若胚不通直,用削尖的铅笔在纸上按实物胚形状画出胚的长度线条,并标出子叶、下胚轴、胚根长度所在位置。用细线绳按纸上画出的线条形状度量,根据拉直的线绳测定胚各部分长度。若胚通直可按实物胚用游标卡尺度量,精度0.01 cm。对无预处理和低温层积种子的离体胚(不包括GA3处理离体胚)以及全胚乳胚和部分胚乳胚的培养,测定了胚萌发生长量。可萌发出胚根的培养处理,在萌发胚中取样;不能萌发出胚根的培养处理,在培养胚中取样。离体胚培养中,按上述取样原则,对萌发胚和培养胚全部测定生长量。带全胚乳胚和部分胚乳胚培养中,各处理均随机取50个胚乳胚测定生长量。
1.2.8 胚萌发能力指标及计算方法采用萌发率Gp和发芽指数Gi 2个指标表示。Gp(%)=胚根萌发总数/培养胚总数×100;Gi(%·d-1)=∑Gt/Dt×100。式中Gt为在时间t日的发芽百分数;Dt为相应的发芽日数。
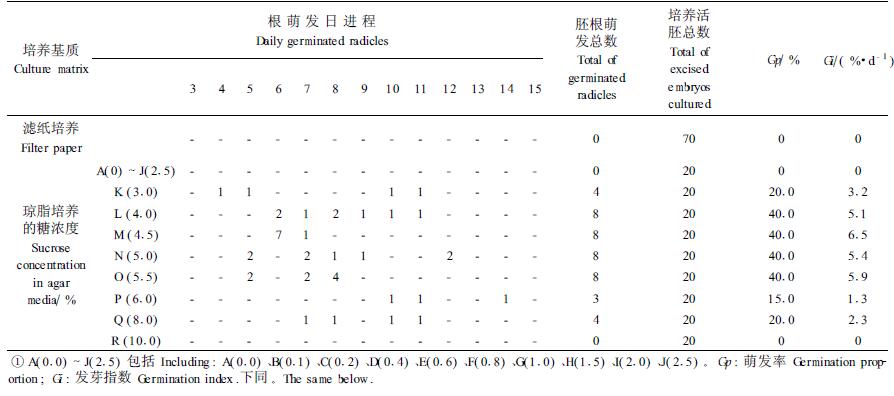
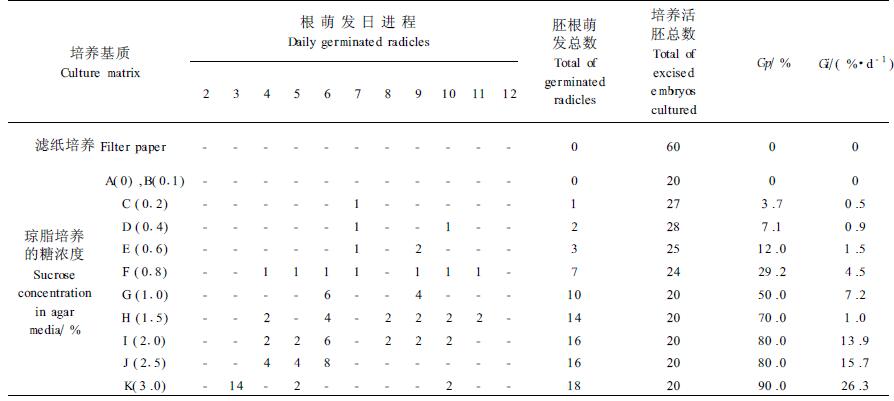
2 试验结果 2.1 无预处理种子离体胚培养无预处理种子离体胚培养在滤纸上及在不加GA3、蔗糖浓度为A(0.0%)~I(2.0%)的琼脂培养基上后,下胚轴就开始伸长、变粗,但无胚根长出。在有蔗糖的琼脂培养基上,胚的下胚轴生长量明显增大(图 1, 图 4)。蔗糖浓度为3.0%的琼脂培养基上培养的20个胚中,于第4、5、10、11天分别有1个胚长出白色幼嫩的胚根。胚根萌发3~4 d后变成棕红色,其余80%的胚始终没有萌发出胚根。未长出胚根的胚在下胚轴的下端有一段红棕色的胚根套,胚根套长0.3~0.9 cm。此培养基上,萌发的胚生长得十分弯曲,几乎折叠成双折(图 2)。在蔗糖浓度为4.0%~5.5%的琼脂培养基上,各有40%的胚在不同时间、以不同的萌发速度(较早的第5天,晚的第12天)萌发出生长良好的白色幼嫩胚根。这些萌发胚不成双折,多数生长成弓形,少数胚根生长较为通直,随着蔗糖浓度增加,通直胚根的比例增大。胚根萌发10 d以后,微带棕色。其余60%的胚始终未萌发出胚根。在蔗糖浓度为6.0%、8.0%、10.0%的琼脂培养基,萌发率下降直至没有胚根萌发出来。离体胚的子叶,在滤纸培养床上和不同蔗糖浓度的琼脂培养基上,培养的第1天部分胚的子叶展开,第2天可全部展开,但生长量极微(表 1)。表 1中胚根萌发数经列联表(χ2)检验,在不同蔗糖浓度培养基上胚根萌发数差异极显著[χ2=52.37>χ0.012(9)=21.67]。

|
图 4 离体胚、部分及全胚乳胚萌发生长量 Fig. 4 Growth of different organs of naked embryos and embryos with partial or intact endosperm 处理1~4为滤纸培养Treatments 1~4 were cultured on filter paper. 1:带全胚乳胚Embryos with intact endosperm(*); 2:带1/2或2/3胚乳胚Embryos with half or more endosperm(*); 3:离体胚Excised embryos(*); 4:离体胚Excised embryos(**).处理5~25为琼脂培养基上离体胚的培养,序号后百分数为蔗糖浓度。Treatments 5~25 were excised embryos cultured on media with different concentrations of sucrose(following No.)respectively.5:0%(**); 6:0%(*); 7:0.1(**);8:0.1%(*);9:0.2%(**);10:0.4%(**);11:0.6%(**);12:0.8%(**);13:1.0%(**);14:1.5%(**);15:2.0%(**);16:2.5%(**);17:3.0%(**);18:3.0%(*);19:4.0%(*);20:4.5%(*);21:5.0%(*);22:5.5%(*);23:6.0%(*);24:8.0%(*);25:10.0%(*)。*:种子无预处理Seeds without pretreatment;**:种子经低温处理Stratified seeds. |
低温层积种子胚比无预处理种子胚置床前长度、粗度大,置床后下胚轴变绿早;萌发率、萌发速度显著提高。在蔗糖浓度3.0%的培养基上,胚培养的第3天就有70%的胚萌发,并且胚根长得十分通直、幼嫩,色白(图 3);在0.2%蔗糖浓度琼脂培养基上已可萌发出胚根。因此经低温层积处理后种子胚的培养,诱导胚根萌发的培养基蔗糖浓度可大幅度降低,萌发率和速度可显著提高。结果见表 2。表 2中胚根萌发数经列联表(χ2)检验差异极显著[χ2=167.78>χ0.012(11)=24.73]。表 1、2中的2种处理离体胚培养萌发率(Gp)及萌发指数(Gi)相比较,经t检验差异均极显著[tGp=3.15>t0.01(26)=2.78;tGi=3.18>t0.01(26)=2.78]。
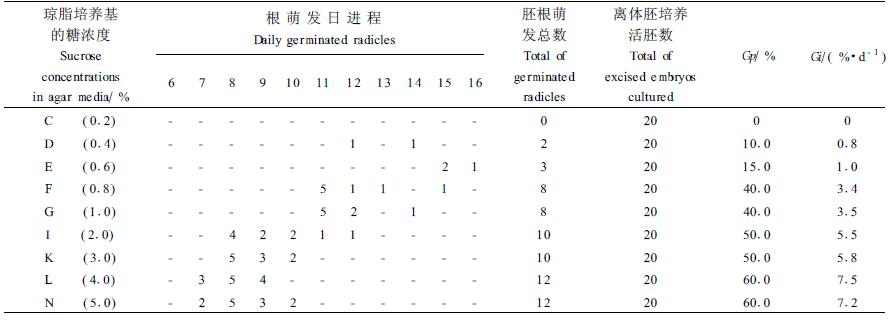
2.3 含GA3培养基离体胚培养含GA3培养基离体胚培养比不含GA3培养基离体胚培养萌发率、萌发速度显著提高,诱导胚根萌发的培养基浓度可大幅度降低(表 3)。表 3中胚根萌发数经列联表(χ2)检验差异极显著[χ2=38.99>χ0.012(8)=20.09]。表 1、表 3中无预处理种子离体胚在具GA3和不具GA3培养基上培养,萌发率(Gp)及萌发指数(Gi)经t检验差异显著至极显著[tGp=3.04>t0.01(24)=2.80;tGi=2.09>t0.05(24)=2.06]。
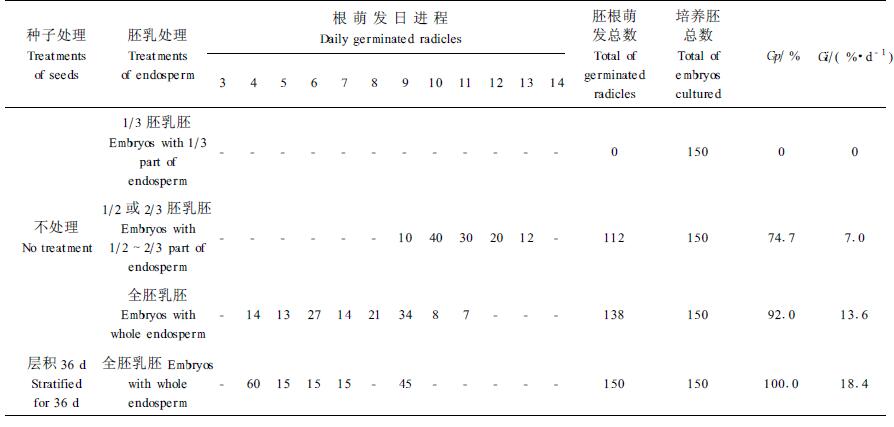
2.4 不同处理带胚乳胚培养带1/3胚乳胚仅下胚轴伸长,不能萌发出胚根。带1/2、2/3、全胚乳胚都可萌发出胚根。但带部分胚乳胚萌发速度慢于全胚乳胚。结果见表 4。表 4中不同处理带胚乳胚胚根萌发数经列联表(χ2)检验差异极显著[χ2=412.90>χ0.012(3)=11.35]。无预处理种子的带全胚乳胚发芽指数(Gi)和带1/2或2/3胚乳胚发芽指数(Gi)比较,经t检验差异极显著[t=3.46>t0.01 (11)=3.11]。
2.5 离体胚、部分及全胚乳胚萌发生长量在滤纸上和只含蔗糖(不同浓度)的琼脂培养基上离体胚子叶生长量增加极少(均可展开)。胚的下胚轴在滤纸上及各种蔗糖浓度琼脂培养基上都可伸长,但蔗糖浓度高的培养基生长量大。能萌发胚根的离体胚若胚根生长量大,下胚轴生长量相对小。离体胚胚根及下胚轴生长量之和,在一定范围随培养基蔗糖浓度的增加而增加。离体胚培养在本试验的各种培养基上以及带部分胚乳胚培养,子叶、下胚轴、胚根生长量远远小于带全胚乳胚培养各器官的生长量。结果见图 4。
3 讨论及结论白皮松种胚具休眠特性无预处理白皮松种子离体胚培养在滤纸或低浓度的蔗糖琼脂培养基上,都不能萌发出胚根来,但当蔗糖浓度增加到3.0%时,部分种胚可萌发出胚根,且在一定范围内随着蔗糖浓度的增加,胚根萌发率及萌发速度也有所增加。这似乎说明白皮松种胚能否萌发,仅仅和蔗糖浓度有关,如果能获取胚萌发所必需的蔗糖条件,胚根就可以萌发,由此似乎可认为白皮松种胚无休眠特性。但若继续增加蔗糖浓度,直至浓度过大,反而抑制胚根萌发,且不能使所有种胚的胚根萌发,这说明胚的萌发能力不仅和蔗糖有关,也和其他因素有关,同时也说明在任何蔗糖浓度下不能萌发出胚根的这部分种胚具休眠特性。那么在适宜蔗糖浓度下可以萌发胚根的种胚是否具休眠特性?根据试验结果分析,这部分胚虽然能萌发,但萌发速度慢、数量少。种子经低温层积后以及培养基中加GA3,胚的萌发率及速度大大提高,诱导胚根萌发的培养基蔗糖浓度大幅降低(表 1~3),说明在适宜蔗糖浓度下能萌发胚根的胚也具休眠特性。这些种胚之所以能萌发出胚根,是因为其休眠程度比较浅,并非不具休眠特性。进一步分析这部分种胚有的萌发快,有的萌发慢,可能是因为休眠深浅不一。同理,适宜蔗糖浓度下始终不能萌发出胚根的种胚是因为休眠程度较深。综上分析,白皮松种胚具休眠特性。
白皮松种胚的下胚轴没有休眠特性白皮松种胚培养在滤纸上以及含多种浓度蔗糖琼脂培养基上,胚的下胚轴都能迅速伸长,说明白皮松种胚的下胚轴没有休眠特性。
白皮松种皮透性不是阻碍胚萌发的主要因素根据表 1~3试验结果可以看出,胚在无内外种皮以及胚乳的包被下并没有解除休眠,休眠仅是在种子经低温层积、GA3处理后才被解除。说明白皮松种皮透性不是阻碍胚萌发的主要因素。这一研究与Jackson(1965)、王文章(1979)、Dong等(1993)的结论一致。
白皮松种胚胚根的萌发需要有足量浓度的蔗糖非低温层积种子、经低温层积种子、GA3处理种胚在滤纸上、无蔗糖及蔗糖浓度很低(各处理要求蔗糖浓度不一致)的琼脂培养基上培养,都不能萌发出胚根,当蔗糖浓度增加到一定浓度后(表 1~3),胚根就可萌发出来,说明白皮松胚根萌发需要有足量浓度的蔗糖。蔗糖在胚萌发中可能起提供碳源、调节培养基渗透压的作用,对胚器官发生可能有重要作用(陈祖铿等,1983)。为什么经低温层积、GA3处理的离体胚能在蔗糖浓度较低的培养基上诱导出胚根(表 1~3)?这可能由于促进萌发的激素能够增强细胞的生理代谢活性,有利于有机物的运输及利用(殷宏章,1983;倪德祥,1987;张继澍,1999)。低温层积(傅家瑞,1985)和GA3处理都会增加离体胚中的GA3及其他促进萌发的激素的含量,增强离体胚生理代谢活性,提高蔗糖利用水平,使离体胚能在蔗糖浓度较低的培养基上诱导出胚根。离体胚在仅含蔗糖的培养基上,子叶生长量极微,这可能是胚的根和苗系统代谢有根本差别,培养基中缺乏与子叶生长有关的物质(陈祖铿等,1983)。
白皮松种胚的萌发受胚乳的调控据表 4的结果可以看出,带全胚乳胚92%(无预处理种子)可萌发出胚根,带1/2或2/3胚乳胚74.7%可萌发出胚根,带1/3胚乳胚只下胚轴伸长,不能萌发出胚根。而且带全胚乳胚萌发速度显著快于带1/2或2/3胚乳胚。这些结果都说明胚乳为胚的萌发可能提供了必需的条件,对胚的萌发具促进作用。但另一方面,经低温层积36 d白皮松种子的离体胚在3.0%蔗糖培养基上有70%的胚在第3天就萌发出胚根,说明这70%的离体胚的萌发能力差异很小,所以能够在同一天萌发,可是同样低温层积36 d的种子进行带全胚乳胚培养,发芽率在4~9 d逐日以40%、10%、10%、10%、0%、30%的萌发速度进行(表 4),说明胚乳对胚的萌发具抑制作用。试验结果还表明这种抑制作用对不同的胚作用程度可能不同,对有的胚作用程度可能较小,这些胚发芽相对较早;对有些胚作用程度可能较大,这些胚发芽相对较晚。据以上分析推测,白皮松种胚的萌发可能受胚乳的调控。这种调控作用,一方面表现在胚乳对胚的萌发具促进作用;另一方面,胚乳对胚的萌发也具抑制作用。
胚乳为什么使已基本解除休眠的胚萌发受阻?这可能是胚乳和胚中所含抑制物的量或性质不同(王文章,1979;董丽芬等,1987),所以种子经低温层积后,胚和胚乳解除休眠程度不一样,胚解除休眠程度可能较大,胚乳相对胚解除休眠程度可能小些,胚乳中的抑制物对胚萌发可能还会起作用的缘故。胚乳对胚萌发的抑制作用为什么对不同胚作用程度又不相同呢?从萌发抑制物的生态学意义(傅家瑞,1985)分析,可能不同胚乳具有和萌发有关的激素水平或激素性质不同,也可能是不同胚乳营养状况不同以及酶的活性不同,也可能是几种原因的综合。胚乳对胚萌发阻碍作用的这种差异性,应该是白皮松种子对外界环境条件的一种适应,是自然选择的结果。
陈祖铿, 母锡金, 孙安慈, 等. 1983. 维管植物实验胚胎发生. 北京: 科学出版社.
|
程淑婉, 程东海, 方炎明. 1992. 松树种子萌发过程中苯丙氨酸解氨酶的活力. 南京林业大学学报, 16(2): 42-46. |
董丽芬, 肖斌, 曹翠萍. 1987. 白皮松种子休眠原因的初步探讨. 陕西林业科技, (1): 15-17. |
傅家瑞. 1985. 种子生理. 北京: 科学出版社.
|
GB 2772-1999.中华人民共和国国家标准林木种子检验规程.1999-11-10发布, 2000-04-01实施
|
黄玉国. 1986. 刺楸种子胚休眠的研究. 东北林业大学学报, 14(1): 39-44. |
李晓洁, 徐化成. 1989. 白皮松种子发芽习性及其种源变异的研究. 林业科学, 25(2): 97-104. DOI:10.3321/j.issn:1001-7488.1989.02.012 |
倪德祥. 1987. 植物生长调节剂在组织培养中的调控作用. 自然杂志, 10(1): 35-39. |
尚杰, 王文章, 赵垦田. 1992. 白皮松种子萌发过程中有机质代谢的研究. 东北林业大学学报, 20(2): 24-29. |
陶章安. 1956. 白皮松等13个树种层积催芽的初步研究. 林业科学, (1): 48-54. |
谭志一, 董愚得, 房耀仁, 等. 1983. 红松种子休眠与脱落酸及外种皮的关系. 中国科学, B辑(9): 816-822. |
王文章. 1979. 红松种子抑制物质的初步研究. 植物生理学报, 5(4): 343-351. |
王文章, 陈杰, 刘恩举. 1986. 红松种子休眠与种皮的关系. 东北林业大学学报, 14(2): 83-86. |
王小平, 刘晶岚, 王九龄, 等. 1998. 白皮松种子及球果形态特征的地理变异. 北京林业大学学报, 20(3): 25-31. DOI:10.3321/j.issn:1000-1522.1998.03.005 |
王小平, 刘晶岚, 王九龄. 2000. 白皮松种子休眠特性的种源变异. 应用生态学报, 11(1): 9-12. DOI:10.3321/j.issn:1001-9332.2000.01.003 |
许绍惠, 韩忠环. 1988. 刺楸种子沙藏过程中胚的发育与休眠. 沈阳农业大学学报, 19(2): 29-34. |
殷宏章(译).生长调节物在农业生产中的应用.北京: 科学出版社, 1983
|
于淑兰. 1983. 解除白皮松种子休眠和促进发芽的试验研究. 种子, (1): 24-27. |
张继澍. 1999. 植物生理学(高等农林院校系列教材). 西安: 世界图书西安出版公司.
|
张良诚, 郭维明, 陈永盛. 1981. 红松种子后熟生理的实验研究. 东北林学院学报, (4): 19-34. |
赵焱, 张学忠, 王孝安. 1995. 白皮松天然林地理分布规律研究. 西北植物学报, 15(2): 161-166. DOI:10.3321/j.issn:1000-4025.1995.02.015 |
Arrillaga I, Marzo T, Seguar J. 1992. Embryo culture of Frainus ornus and Sorbus domestica remove seed dormancy. Hort Scince, 27(4): 371. DOI:10.21273/HORTSCI.27.4.371 |
Asakawa S. 1961. A preliminary study concerning the relationship between an embryo and endosperm during stratification. J Japan Forest Society, 43(6): 195-197. |
Barnett J P. 1972. Seedcoat influences dormancy of Loblolly Pine seeds. Canada Journal of Forest Research, 2(1): 7-10. DOI:10.1139/x72-002 |
Baron F J. 1966. Embryo growth and seed germination of Sugar Pine (Pinus lambertiana Dougl.). Advanc Front Plant Science, New Delhi, 17: 1-13. |
Chapman R. 2000. Embryo and seed coat factors produce seed dormancy in capeweed (Arctotheca calendula). Australian journal of agricultural research, 51(7): 849-854. DOI:10.1071/AR00050 |
Dawidowica Grgegorgewska A, Beranger novat N. 1989. Ultrastructural studies on development in mature seed embryos of Euonymus europae L. dormant on cultures at +25℃. Journal of Experimental Botany, 40(217): 913-918. |
Dong L F, Shao C B, liu G Z et al. Determination of the major cause of seed dormancy of Pinus bungeana Zucc. Dormancy and barriers to germination. Proceedings of an international symposium of IUFRO Project Group P2.04-00(Seed Problems), Forestry Canada, Pacific Forestry Centre, 1993: 31-36
|
Goo M, Negisi K. 1952. Changes of reserve foods in seeds of Kuromatsu (Pinus thunbergii Parl.) during the course of germination. Bulletin Tokyo University Forestry, 43: 83-89. |
Gurt A, Urban B, Lennart B, et al. 1998. Predicting germination capacity of Pinus sylvestris and Picea abies seeds using temperature data from weather stations. Candian Journal of Forest Research, 28(10): 1530-1535. DOI:10.1139/x98-139 |
Hellum A K, Hackett L. 1988. Variable dormancy in seed of Pinus contorta. Scandinavian Journal of Forest Research, 3(2): 137-146. |
Jackson G A D, Blundell J B. 1965. Germination of Rosa arvensis. Nature, 205: 518-219. DOI:10.1038/205518a0 |
Jordy M N, Danti S, Miched J, et al. 2000. Biochemical Changes in Pinus spp. seeds during germination and post-germinative growth: triacylglycerol distribution and activity. Functional plant Biology, 27(12): 1117. |
Kao C. 1972. The mechanism of stratification in the seed of Pinus radiata D. Don. Technical Bulletin Experimental Forest of National Taiwan University: 99-177. |
Kao C. 1970. The excised embryos culture of Pinus morrisonicola(P. parviflora) and Pinus armandii. Quarterly Journal Chinese Forestry, Taipei, 3(2): 129-133. |
Lammer D L, Gifford D J. 1989. Lodgepole pine seed germination. Ⅱ. The seed proteins and their mobiligation in the megagametophyte and embryonic axis. Canada Journal of Botany, 67(9): 2544-2551. DOI:10.1139/b89-329 |
Le P D M T. 1977. Non-Acidic inhibitors and embryo dormancy in Taxus baccata. Physiologia-Plantarum, 41(1): 85-88. |
Le P D M T. 1973a. An embryo-culture study of embryo dormancy in Taxus baccata. Biologica-Plantarum, 15(4): 264-269. DOI:10.1007/BF02922707 |
Le P D M T, Garello G. 1973b. Embryo dormancy in Taxus baccata: influence of culture medium on initiation of germination. Physiologia-Plantarum, 29(2): 204-207. DOI:10.1111/j.1399-3054.1973.tb03093.x |
Sambeek J W V, Preece J E, Lindsay T L et al. In vitro studies on black walnut embryo dormancy. 80th Annual Report-Northern Nut Grower's Association, 1989, Publ., 1990: 50-59
|
Shen T Y, Odén P C. 1999. Activity of sucrose synthase, soluble acid invertase and fumarase in germinating seeds of Scots Pine (Pinus sylvestris L.) of different quality. Seed science & technology, 27(3): 825-838. |
Siti N, Hidayati M, Baskin. 2000. Dormancy-breaking and germination requirements for seeds of Diervilla lonicera(Caprifoliaceae), a species with underdeveloped linear embryos. Canadian Journal of Botany, 78(9): 1199-1205. DOI:10.1139/b00-094 |
Stone E C. 1957a. Embryo dormancy of Pinus jeffreyi Murr. seed as affected by temperature, water uptake, stratification and seed coat. Plant Physiology, 31(2): 93-99. |
Stone E C. 1957b. Embryo dormancy and embryo vigor of Sugar Pine (Pinus lambertiana) as affected by length of storage and storage temperatures. Forest Science, 3(4): 357-371. |
Widrlechner M P, Kovach D A. 2000. Dormancy breaking protocols for Cuphea seed. Seed Science & Technology, 28(1): 11-27. |