 2003, Vol. 39
2003, Vol. 39文章信息
- 张焕朝, 徐成凯, 王改萍, 徐锡增, 胡正义.
- Zhang Huanchao, Xu Chengkai, Wang Gaiping, Xu Xizeng, Hu Zhengyi.
- 杨树无性系根系吸收H2PO4-动力学特征与磷营养效率
- PHOSPHATE UPTAKE CHARACTERISTICS OF KINETICS AND PHOSPHORUS EFFICIENCY IN CLONES OF POPLAR
- 林业科学, 2003, 39(6): 40-46.
- Scientia Silvae Sinicae, 2003, 39(6): 40-46.
-
文章历史
- 收稿日期:2001-12-11
-
作者相关文章


2. 中国科学院南京土壤研究所 南京 210008
2. The Institute of Soil Science, Chinese Academy of Sciences Nanjing 210008
磷是植物生长发育所必需的主要营养元素之一。土壤缺磷是植物高产的重要限制因子。传统上,常用施磷肥来改善植物的磷营养,但是,由于磷素在土壤中易被固定导致利用率很低以及大量使用磷肥带来的诸如资源匮乏、能源危机和环境污染,使得这一传统的农业技术措施面临重重困难。因此,应用生物技术,选择、培育和利用磷营养高效型植物是改善缺磷状况的一条节能、低耗、高产、高效的新途径。而在产生植物磷营养效率差异的诸多因素中,植物根系磷吸收动力学特性具有特殊的作用和意义。
植物之间或同种植物的不同品种之间,根系的吸收动力学特性不同,这说明植物根系的吸收动力学特性具有遗传性。许多研究结果表明根系吸收动力学特性是植物效率差异的一个重要因素,不同基因型植物根系对营养离子的吸收特性具有较大的差异(Claassen et al., 1974;Drew et al., 1984;Jungk, 1974; Jungk et al., 1975;Nielsen et al., 1978;1983)。植物根系吸收营养离子的动力学特性的特征主要是通过吸收动力学参数来描述的。这些动力学参数包括最大吸收速率Vmax,米氏常数Km和吸收临界浓度Cmin。Km是吸收速率为最大吸收速率的一半时外界离子浓度,1/Km表征吸收系统的亲合力,Km值小,说明根系吸收系统对该离子的亲合能力大;Cmin是净吸收为0时外界离子的最低浓度,Cmin值小,表明植物能从有效性非常低的土壤中吸收该养分,对低养分的耐受能力强。国内外对农作物的根系吸收动力学特性进行了大量研究。Nielsen等(1978;1983)对玉米和大麦的研究表明,不同磷营养效率的基因型植物的Km和Cmin不同,磷营养效率高的基因型有Vmax大,Km和Cmin小的特点。林翠兰等(1992)对玉米不同品种的根系吸收动力学特性的研究也得出类似结论。近年来,国外对林木,尤其是苗木根系吸收动力学特性的研究逐渐增多。Kelly等(1991;2001a;2001b)分别对火炬松(Pinus taeda)、红花槭(Acer rubrum)苗和一些阔叶树苗进行了根系磷、钾和镁的吸收动力学特性研究;Pedersen等(1999)发现与草本植物(Panicum chamaelonche)相比,湿地松(Pinus elliottii)苗具有较高的Vmax和Cmin。国内在这方面的报道较少,仅见方从兵等(2001)对毛桃(Prunus persica)幼苗磷、钾吸收动力学的研究。本研究选择我国主要速生造林树种杨树为对象,研究其根系吸收H2PO4-的动力学特性,探讨其与杨树无性系磷营养效率的关系,以揭示杨树无性系磷营养效率差异的机理,并为杨树无性系新品种的培育提供理论依据和积累科学资料。
1 材料与方法 1.1 试验材料采用美洲黑杨(Populus deltoides)与四川青杨(P. cathayana)、小叶杨(P. simonii)、欧美杨(P. euramericana)3个杂交组合F1代中选育出的6个新无性系和2个美洲黑杨无性系共8个无性系作试材,插穗长25 cm,粗2~2.5 cm。无性系分别为:美洲黑杨×四川青杨(P. deltoides×P.cathayana)F1(简称S17);美洲黑杨×四川青杨(P. deltoides×P.cathayana)F1(简称S19);美洲黑杨×小叶杨(P. deltoides×P.simonii)F1(简称105);美洲黑杨×小叶杨(P. deltoides×P.simonii)F1(简称106);美洲黑杨×欧美杨(P. deltoides×P.euramericana)F1(简称797);美洲黑杨×欧美杨(P. deltoides×P.euramericana)F1(简称1388);美洲黑杨Ⅰ-69 (P. deltoides cv. `Lux')(简称69);美洲黑杨3244 (P. deltoides cv.3244)(简称3244)。
1.2 试验方法植物根系吸收动力学特性的研究通常有2种方法:1.切离根的短期吸收(short term absorption by excised roots);2.整株植物长期吸收(long term absorption by intact plants)。第1种方法由于只能用于同位素标记的离子的吸收特性测定,并且吸收阶段较短,奈等(1985)认为这种方法的测定结果不能确切地反映根系实际吸收状况。第2种方法似乎更符合实际吸收情况,且应用范围更广泛。本研究应用第2种方法,即整株植物长期吸收的培养方法,分2个阶段,即培养阶段和吸收试验阶段。
1.2.1 培养阶段将无性系插穗固定在盛有2 L相应处理的营养液的培养桶中,每桶2株。试验设2个磷水平(+P与-P),+P为施磷处理,-P为不施磷处理。+P施磷处理营养液的组成参照Claassen等(1974)和林翠兰等(1992)推荐的配方,并适当调整为:CaSO4 0.5 mmol·L-1, KNO3 0.75 mmol·L-1, Mg(NO3)2 0.35 mmol·L-1, KH2PO4 0.5 mmol·L-1, Fe-EDTA 1×10-4 mol·L-1, H3BO4 8×10-6 mol·L-1, CuSO4 1×10-6 mol·L-1, ZnSO4 2×10-7 mol·L-1, (NH4)6MoO24 2×10-7 mol·L-1, MnSO4 2×10-7 mol·L-1。-P处理的营养液组成是将上述+P处理营养液中0.5 mmol·L-1的KH2PO4用0.5 mmol·L-1的KCl代替,其余组分不变。培养液的pH值调节至6.0±0.1,光照为20 klx,培养温度为(25±2~15±2)℃。每2 d换一次培养液,连续通气。培养时间为4月15日至5月15日。培养液体积为2 L,3次重复。培养试验在温室中完成。
1.2.2 吸收试验阶段采用Claassen等(1974)提出的离子消耗曲线方法(the depletion curve procedure)研究整株杨树无性系根系对离子的吸收与外界离子浓度的关系。该方法的一个最基本的假设就是一般认为,在低浓度范围内,根系对许多离子的吸收服从Michaelis-Menten动力学方程。因此,可以用Michaelis-Menten动力学方程描述根系对离子的吸收状况。该方法的优点在于:1.采用整株植物在同一培养容器中连续吸收培养,可以准确地描述吸收速率与培养液浓度之间的关系,减少了因植株之间差异所造成的实验误差;2.可以得到由Vmax、Km和Cmin表达的连续函数,减少了环境因素如温度、植株年龄、光照强度等对离子吸收的影响;3.可以估计植物吸收离子的临界浓度Cmin参数;4.比较适合于H2PO4-、K+的吸收动力学研究。5月15日,将上述培养阶段的无性系试验材料转入去离子水中,让植株处于饥饿状态,保持24 h。然后转入2 L含有0.05 mmol·L-1 KH2PO4的营养液中(其它营养成分的浓度均为原培养阶段组分的1/10)。分别在0.5,1,2,2.5,5,8,12,13,14 h后取样测定吸收液中H2PO4-的浓度。各处理每次取样2 mL,同时补充2 mL去离子水。吸收试验期间,温度为(25±2)℃,光照为20 klx,连续通气,保持吸收液的pH值在6.0±0.1。试验重复2次。
1.2.3 吸收液H2PO4-离子浓度的测定用钼蓝法测定吸收液中P的浓度,再换算为H2PO4-离子的浓度,μmol·L-1。
2 结果与分析 2.1 杨树无性系对H2PO4-的吸收状况 2.1.1 -P处理下无性系对H2PO4-的吸收经-P处理的杨树无性系,在低浓度全营养液里生长,随吸收时间的延长,培养液中H2PO4-的浓度逐渐降低,但下降的幅度不同,无性系S17、S19和105培养液中H2PO4-的下降幅度明显大于无性系106、797、Ⅰ-69、1388和3244(表 1)。由于无性系S17、S19和105是磷营养高效型无性系,而106、797、Ⅰ-69、1388和3244均为磷营养低效型无性系(张焕朝等,2001),因此经-P处理后高效型无性系对H2PO4-的吸收能力大于低效型无性系。例如经过8 h的吸收,高效型无性系S17、S19和105培养液中H2PO4-浓度分别从50 μmol·L-1下降到3.25、4.06和4.92 μmol·L-1,而低效型无性系溶液浓度下降较少,培养液中H2PO4-浓度均在17 μmol·L-1以上。
|
|
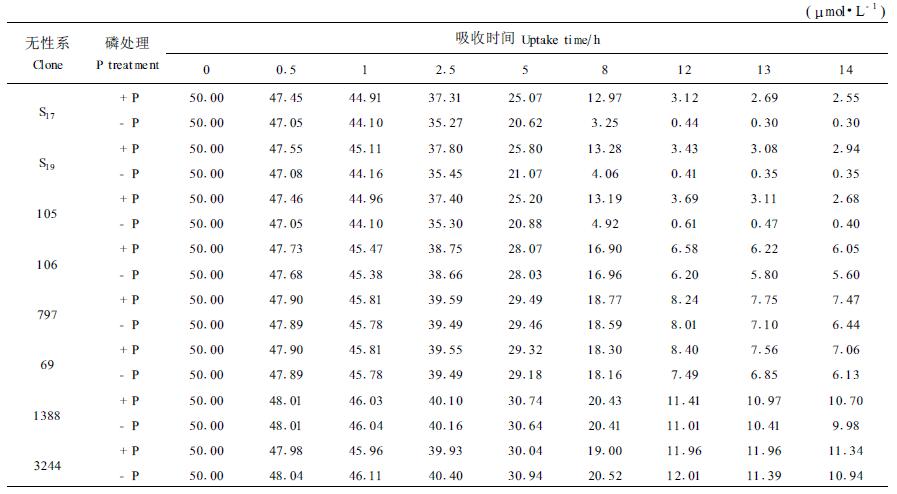
同-P处理类似,+P处理的杨树无性系再次放入低浓度全营养液里生长,培养液中H2PO4-的浓度也随吸收时间的延长而下降,但下降幅度有差异。降幅最快的也是S17、S19和105三种高效型无性系(表 1),说明供磷充足时,高效型无性系对H2PO4-的吸收能力也较强,而低效型无性系对H2PO4-的吸收能力较弱,表现在培养液中H2PO4-的浓度下降幅度较小。如1388和3244经12 h吸收后,培养液浓度仍为12 μmol·L-1左右,而高效型无性系则已降低到4 μmol·L-1以下。
2.1.3 磷营养状况对无性系根系吸收H2PO4-的影响无性系对H2PO4-离子的吸收动态还反映了不同供磷状况下各无性系对H2PO4-吸收能力的差异。从0~14 h,-P处理的高效型无性系培养液中H2PO4-的浓度下降幅度均明显地高于同一种无性系的+P处理。例如,S17吸收5 h时培养液浓度经-P处理仅20.62 μmol·L-1,而+P处理却为25.07 μmol·L-1(表 1),表明高效型无性系在缺磷胁迫后对H2PO4-的吸收能力增强。也就是说,高效型无性系可以通过增加根系的吸收能力对低磷逆境作出适应性反应。而低效型无性系不具有这种适应性反应。
从表 1还可以看出,无论是经+P处理,还是经-P处理,随着吸收时间的延长,各无性系的离子消耗曲线逐渐趋于平滑,特别是在吸收时间达到13 h和14 h后。此时培养液中H2PO4-浓度已降低到一个很低的程度,基本上不再降低或降低很小。这表明无性系在此时从外界溶液吸入离子的量(influx)与其根系向外界渗出的量(efflux)达到平衡,即净吸收为0。
2.2 杨树无性系根系对H2PO4-的吸收动力学特征 2.2.1 无性系吸收动力学参数根据各无性系对H2PO4-的消耗曲线,可以获得各无性系在不同吸收培养液浓度时的吸收速率。吸收速率为无性系单位时间单位根干重所吸收的H2PO4-的量。它实际上是离子消耗曲线的导数,如果令Q=f(t)表示离子消耗曲线函数,则吸收速率V=-dQ/dt。以V-C作图,可得各无性系吸收速率和培养液浓度的关系曲线,如图 1所示。杨树无性系根系对H2PO4-的吸收速率(V)随培养液浓度(C)的增加而增加(图 1)。当培养液浓度增加到某一值时,吸收速率达最大,此时吸收速率为Vmax。各无性系的V-C曲线基本上服从于Michaelis-Menten动力学方程,即V=Vmax·C/(Km+C)。由于植物根系在吸收养分的同时也发生该养分的外流或渗出,特别是磷、钾,因此,用净吸收速率来描述根系对离子的吸收比较合理,所以上述方程应修正为:V=Vmax·(C-Cmin)/(Km+C-Cmin)。式中,V为某浓度时的净吸收速率,Vmax为最大吸收速率,Km为米氏常数(Michaelis-Menten常数),C为外界溶液浓度,Cmin为吸收临界浓度。

|
图 1 杨树无性系根系对P的吸收速率与培养液H2PO4-浓度的关系 Fig. 1 Relation between the rates of P uptake by root systems of poplar clones and H2PO4- concentration in external solution under various P treatments |
由图 1可得能够描述各无性系根系动力学吸收特性的参数Vmax、Km和Cmin。如表 2所示。关于根系动力学吸收参数Vmax、Km和Cmin的获得,不同的研究者采用不同的方法。Claassen和Barber (1974)将Michaelis-Menten动力学方程与离子消耗曲线数据拟合,再通过计算获得。林翠兰等(1992)通过查阅曲线获得。本研究的Vmax、Km和Cmin是通过曲线查阅得到。
|
|
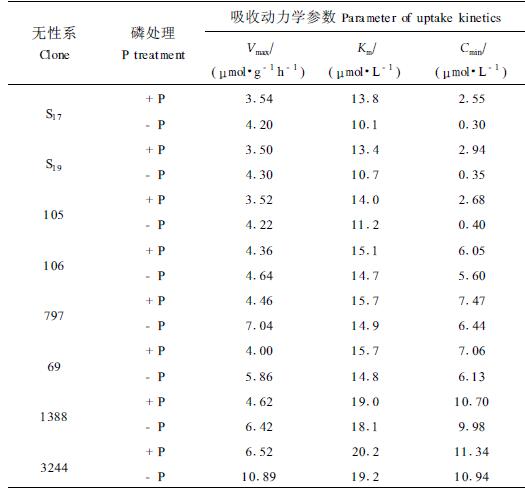
经-P处理后,无性系S17、S19和105的Vmax约在4.20~4.30 μmol·g-1h-1之间,而106、797、69、1388和3244则在4.00~10.89 μmol·g-1h -1之间(表 2)。高效型无性系的Vmax比低效型无性系低。这与通常所认为磷营养高效基因型植物具有较高的最大吸收速率不一致。事实上,有不少研究者认为最大吸收速率在指示高磷供应对植物生长的限制上更有意义(Anghinoni et al., 1980;Barber,1984;Borkert et al., 1985)。
吸收常数Km (米氏常数)高效型无性系则明显低于低效型无性系。S17、S19和105的Km值分别为10.1、10.7和11.2 μmol·L-1, 而无性系106、797和69则均在14.5 μmol·L-1以上,1388和3244更是接近20 μmol·L-1(表 2)。由于吸收常数的大小反映了根系系统对H2PO4-的亲合能力,是具有特征性的指标,Km越小,说明亲合力越强,因此高效型无性系对H2PO4-离子的亲合能力明显高于低效型无性系。如S17的1/Km值大约是3244的1.9倍以上。甚至亲合能力差别最小的也在30%以上,如105的亲合力约高出106无性系31%。
根系对H2PO4-的吸收临界浓度Cmin反映了无性系对低磷的耐受能力。Cmin越小,说明该无性系越能从有效磷低的环境中吸收磷,从而对低磷的耐受能力就越强。经-P处理后,无性系S17、S19和105的Cmin值均在0.40 μmol·L-1以下,大大低于磷营养低效型无性系106、797、69、1388和3244。后者的Cmin值均在5.50 μmol·L-1以上,尤其是1388和3244,Cmin值达10.00 μmol·L-1(表 2),这意味着当H2PO4-离子的浓度尚有10 μmol·L-1之丰时,它们已经基本上不能吸收这些磷素了。高效型无性系(如S17、S19和105)对低磷的耐受程度要比低效型无性系高出14~25倍。强大的耐受低磷能力可能是S17、S19和105等无性系之所以磷营养效率高的重要原因之一。
2.2.3 +P处理条件下无性系根系吸收动力学特性的差异经+P处理后,最大吸收速率Vmax同样为高效型无性系小于低效型无性系。米氏常数Km,高效型无性系在13.4~14.0 μmol·L-1之间,而低效型无性系在15.1~20.2 μmol·L-1之间。虽然仍表现出高效型无性系对H2PO4-离子亲合能力强于低效型无性系这一趋势,但差异程度明显下降,除1388和3244仍相差较大外,其它无性系之间的差异很小(表 2)。这可能是在供磷充足的环境条件下,由于根系对离子的主动吸收比例相对减少,被动吸收的比例增大,从而抑制了根系对H2PO4-的亲合能力。
高效型无性系的根系吸收临界浓度Cmin小于低效型无性系。前者在2.55~2.94 μmol·L-1之间,后者在6.05~11.34 μmol·L-1之间。说明高效型无性系的耐受低磷能力在供磷充足的环境中仍强于低效型无性系,尽管这种耐受低磷能力的差异程度因磷供应充足而减小了。
2.2.4 磷营养状况对无性系根系吸收动力学特性的影响磷营养状况对无性系的最大吸收速率、吸收常数和吸收临界浓度会产生较大影响。与+P处理相比,各无性系的Vmax经-P处理后均有增加。高效型无性系增加20%左右,而低效型无性系增加较多,约在45%~65%之间(表 2)。吸收常数Km对磷营养状况的反应,不同无性系表现不一。缺磷胁迫使S17、S19和105三种无性系的Km分别降低26.8%、20.1%和20.0%,降幅均在20%以上。表明其根系吸收系统对H2PO4-的亲合力在受到磷胁迫时明显增强。而低效型无性系106、797、69、1388和3244的Km的降幅很小,均在6%以下(表 2)。说明低效型无性系的根系吸收系统对H2PO4-的亲合力基本上不能根据环境中磷的供应状况作出主动反应。这一研究结果与其他对农作物的研究结论基本一致(Nielsen et al., 1978;1983;林翠兰等,1992),证明磷营养高效型无性系能在缺磷胁迫条件下,通过某种机制主动改变其亲合能力,从而达到磷营养高效的目的。各无性系根系吸收H2PO4-的临界浓度Cmin也受到磷营养状况的影响。S17、S19和105经-P处理后Cmin分别降低了88.2%、88.1%和85.1%,表明高效型无性系受低磷胁迫时对低磷的忍耐能力可提高85%以上。而低效型无性系在不同磷营养环境中Cmin的差异则很小,其降幅均在15%以下。
3 结论和讨论杨树无性系在根系吸收H2PO4-动力学特性方面存在着较大的差异。高效型无性系S17、S19、105具有对H2PO4-亲合力强,耐低磷能力强的特点,表现在其具有较小的Km和Cmin值,并且Km和Cmin在缺磷胁迫的环境中有大幅度的下降。这说明磷营养高效型无性系可以通过某种机制对缺磷胁迫作出主动反应,增强其根系系统对H2PO4-的亲合能力和提高其对低磷的耐受能力,从而使它们具有较高的磷营养效率,表现为在缺磷的环境中有较好的生长。而低效型无性系的Km和Cmin值较大,说明对H2PO4-的亲合能力和忍受低磷能力均较差,并且在缺磷环境中不具有主动改变其亲合能力和耐受能力的吸收动力学特性。根系吸收常数Km和吸收临界浓度Cmin作为描述根系吸收动力学特性的重要参数,在研究杨树无性系磷营养效率差异的机理方面有着非常重要的意义,较低的Km和Cmin是无性系S17、S19和105成为磷营养高效型无性系的重要原因。
至于最大吸收速率Vmax,虽然有研究者认为磷营养高效植物基因型具有较大的Vmax,但也有不少研究者认为Vmax在指示高磷状况下增产所受到的限制更有意义。本研究发现高效型无性系的Vmax实际上并不比低效型无性系大,甚至低于低效型无性系。这一方面说明Vmax作为根系吸收动力学参数之一,在阐明营养效率高、低这一问题时的作用尚不够确定;另一方面,在表示Vmax时所用的单位也应谨慎。考虑到林业研究的对象大都是高大的乔木,即使在苗期也具有比农作物大得多的根系干重,因此用根系干重表示Vmax可能会在一定程度上掩盖其真实差异。尽管国内外不少研究者在对农作物做的类似研究中都指出可以用根系干重代替根系长度或根表面积来表示Vmax,但林业上用根系长度或根表面积来表示Vmax可能会取得更好的结果,这在今后的研究中应加以考虑。
方从兵, 孙其宝, 孙俊. 2001. 桃K+、H2PO4-吸收动力学研究. 中国林副特产, 58(3): 4-5. DOI:10.3969/j.issn.1001-6902.2001.03.003 |
林翠兰, 曹一平, 张福锁.缺磷对不同基因型玉米根系形态及吸收生理特性的影响.见: 张福锁主编.土壤与植物营养新动态.第一卷.北京: 北京农业大学出版社, 1992: 120-124
|
奈P H, 廷克P B著.夏荣基等译.溶质在土壤-根系系统中的运动.北京: 科学出版社, 1985: 124-126
|
张焕朝, 徐成凯, 王改萍, 等. 2001. 杨树无性系的磷营养效率差异. 南京林业大学学报, 25(2): 14-18. |
Anghinoni I, Barber S A. 1980. Phosphorus influx and growth characteristics of corn roots as influenced by phosphorus supply. Agron J, 72: 685-688. DOI:10.2134/agronj1980.00021962007200040028x |
Barber S A. 1984. Soil nutrient bioavailability:A mechanistic approach. New York: John Wiley and Sons.
|
Borkert C M, Barber S A. 1985. Soybean shoot and root growth and phosphorus concentration as affected by phosphorus placement. Soil Sci Soc Am J, 49: 152-155. DOI:10.2136/sssaj1985.03615995004900010031x |
Claassen N, Barber S A. 1974. A method for characterizing the relation between nutrient concentration and flux into roots of intact plants. Plant Physiol, 54: 564-568. DOI:10.1104/pp.54.4.564 |
Drew M C, Saker L R. 1984. Uptake and long-distance transport of phosphate, potassium, and chloride in relation to internal ion concentration in barley: evidence of nonallosteric regulation. Planta, 160: 500-507. DOI:10.1007/BF00411137 |
Jungk A. Phosphate uptake characteristics of intact root systems in nutrient solution as affected by plant species, age and P supply. Hannover: Proc. 7th Intern. Coll. on Plant Analysis and Fertilizer Problems, 1974
|
Jungk A, Barber S A. 1975. Plant age and uptake characteristics of trimmed and untrimmed corn root systems. Plant Soil, 42: 227-239. DOI:10.1007/BF02186985 |
Kelly J M, Barber S A. 1991. Magnesium uptake kinetics in loblolly pine seedlings. Plant Soil, 134: 227-232. DOI:10.1007/BF00012040 |
Kelly J M, Kelly J K. 2001a. Phosphorus and potassium uptake kinetics in red maple seedlings. Forest Science, 47: 397-402. |
Kelly J M, Scarborough J D, Mays P A. 2001b. Hardwood seedling root and nutrient parameters for a model of nutrient uptake. J Environ Qual, 30: 427-439. DOI:10.2134/jeq2001.302427x |
Nielsen N E, Barber S A. 1978. Differences among genotypes of corn in the kinetics of P uptake. Agron J, 70: 695-698. DOI:10.2134/agronj1978.00021962007000050001x |
Nielsen N E, Schjerring J K. 1983. Efficiency and kinetics of phosphorus uptake from soil by various barley genotypes. Plant Soil, 72: 225-230. DOI:10.1007/BF02181961 |
Pedersen C T, Silvia D M, Shiling D G. 1999. Piolithus arhizus ectomycorrhiza affects plant competition for phosphorus between Pinus elliottii and Pacicum chamaelonche. Mycorrhiza, 9: 199-204. DOI:10.1007/s005720050267 |