 2003, Vol. 39
2003, Vol. 39文章信息
- 徐庆, 姜春前, 郭泉水, 刘世荣, 郝玉光.
- Xu Qing, Jiang Chunqian, Guo Quanshui, Liu Shirong, Hao Yuguang.
- 濒危植物四合木结实特性与植株年龄和生境关系的研究
- A STUDY ON THE RELATIONSHIP BETWEEN FRUITING CHARACTERISTICS AND REPRODUCTIVE AGE, HABITAT OF ENDANGERED SPECIES TETRAENA MONGOLICA
- 林业科学, 2003, 39(6): 26-32.
- Scientia Silvae Sinicae, 2003, 39(6): 26-32.
-
文章历史
- 收稿日期:2002-12-04
-
作者相关文章


2. 中国林业科学研究院林业研究所 北京 100091;
3. 中国林业科学研究院沙漠化林业实验中心 磴口 015200
2. The Research Institute of Forestry, CAF Beijing 100091;
3. Experiment Center of Desert Forestry of CAF Dengkou 015200
四合木(Tetraena mongolica)为蒺藜科(Zygophyllaceae)中一个单种属的古老残遗物种,属强旱生灌木。主要分布于鄂尔多斯高原西北部、库布齐沙漠以南、阿拉巴素山的山麓地带。它的存在不仅对认识古地中海植物区系和植被起源具有重要的理论意义,而且对我国西部地区生态环境保护也具有很大的价值。近年来,由于多种因素影响,其分布面积和个体数量逐渐减少,而被列为国家二级重点保护植物。
植物生活史中以种子形态出现的阶段,被称为植物的潜在种群(Harper, 1977; Silvertown, 1982; Steven, 1991)。种子阶段是有性繁殖的植物个体一生中唯一有移动力的阶段,因此对于植物种群的分布格局、种群动态及种群的调控等方面均有重要意义(Harper, 1977; Steven, 1991; 谢宗强, 1998)。
四合木主要通过种子繁殖进行更新。在自然群落中发现,四合木植株虽生长良好,看上去果实累累,但实际上果皮内饱满的种子却寥寥无几。近年来,吴素琴等(1994)在实验室内对四合木5项种子特性即千粒重、吸水增重率、发芽率、存活力、生活力进行了测定,徐庆等(2001)对其种群的生殖值、生殖分配进行了研究报道,王迎春等(2001)对其种群生殖对策进行了探讨。有关四合木种群的结实规律与植株年龄和生境关系的研究未见报道。本文从生殖生态学角度对四合木种群的初次结实年龄,果实的空间分布、果实的数量、果实的重量、结籽率及花果转移率随年龄和生境的变化规律进行研究,并对结籽率低的原因进行了分析,以期为更深入揭示四合木种群的濒危机制提供科学理论依据。
1 研究地区概况及研究方法 1.1 自然概况所选样地位于内蒙古杭锦旗丘地及滩地,位于北纬39°45′~40°20′,东经106°40′~107°20′。本区大陆性气候明显:冬季寒冷,风大沙多;夏季酷热,干旱少雨,热量丰富。全年日照总时数平均为2 792.2 h,年均温9.7℃,极端最高气温达39℃,极端最低气温达-32℃;地面最高温度达63.4℃,地面最低温度为-23.4℃。年平均降水量137.8 mm,大都集中在6—8月;年均蒸发量3 217.7 mm,干燥度4.05。
杭锦旗丘地的植物群落以四合木为优势种,上层有红沙(Reaumuria soongorica)、狭叶锦鸡儿(Caragana stenophylla),其下有珍珠柴(Salsola passerina)、驼绒藜(Ceratoides latens)伴生。多年生丛生禾草形成亚优势层片,主要代表有无芒隐子草(Cleistogenes songarica)、沙生针茅(S. glareosa)、戈壁针茅(S. gobica)、短花针茅(S. breviflora)等。
杭锦旗滩地的植物群落以霸王柴(Zygophyllum xanthoxylon)、四合木为优势种,并与少量沙冬青(Ammopiptanthus mongolicus)构成群落的上层;其下有绵刺(Potaninia mongolica)、刺叶柄棘豆(Oxytropis aciphy)等小灌木;无芒隐子草、沙生针茅、戈壁针茅等多年生禾草在下层形成亚优势层片;其它草本成分有戈壁天门冬(Asparagus gobicus)、黄蒿(Artemisia scoparia)、草霸王(Zygophyllum mucronatum)、阿氏旋花(Convolvulus ammannii)、蒙古葱(Allium monglicum)和多根葱(A. polyrrhizum)等。群落及生境特征见表 1。
|
|
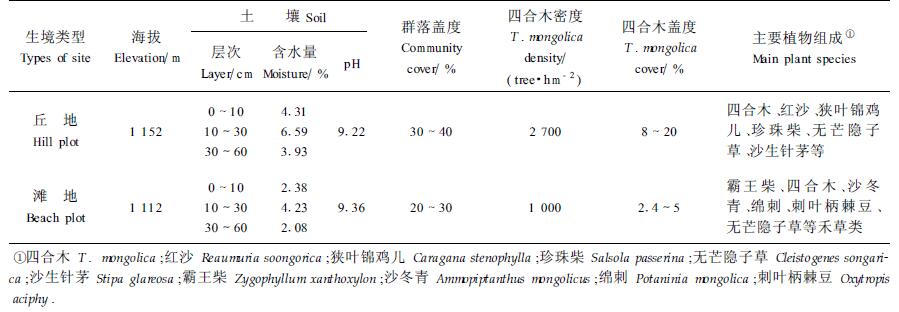
根据群落学调查方法,在杭锦旗丘地及滩地,各设置宽度为10 m、长度为3 800 m样带一个,在样带上每隔100 m设置1个10 m×10 m的样方。每个样带共设置35个样方。在样方内精确测定每株四合木的地径和冠幅(精确度为0.10),同时调查群落植物组成,统计四合木的株数。在每条样带上设置5个土壤采集点,按0~10,10~30和30~60 cm 3个层次分别取样,分析测定土壤含水量、pH等,调查样地群落及生境特点(表 1)。
随机截取77株四合木标准植株基径样品,做好标记,带回实验室进行切面、磨光,用瑞典产的年轮分析仪(CXDDO型)查数年轮,建立年龄(A)与地径(D)的回归方程:A =0.14+0.670 9D,相关系数R=0.918 86(n=77,p<0.05)。以此方程为基础,计算调查地区每株四合木的年龄。
在丘地和滩地,以3 a为1个龄级对四合木种群进行龄级划分,按每龄级5株,各选45株四合木植株,在5月份花蕾期及7—8月份果实脱落前,查数这些植株的花蕾数和初果数,计算花果转移率(花果转移率=果实数/花蕾数)。根据所调查的种子数、果实数,分别计算每株四合木结籽率(结籽率=种子数/胚珠数×100%,胚珠数=果实数×4)。
在四合木开花前(花蕾期),随机选取8株个体,整个花序套尼龙网(网眼大小为1 mm×1 mm, 完全隔离了昆虫传粉),去雄不套袋、不去雄套袋实验,最后统计结籽率,分析结籽率低的原因。
在丘地和滩地,以3 a为一龄级对四合木种群进行龄级划分。每个龄级选择5株四合木植株,分别测量其地径。区分南北、东西2个方向,上、中、下3个层次6个部位,在果实脱落前,查数果实数和种子数。并取部分果实样品,置烘箱内烘至恒重,称其果实的干重,通过对取样数据的统计分析,得出四合木果实的空间分布规律,各龄级平均产果实数、干重及结籽率,及其与年龄的关系。
2 结果与分析 2.1 果实空间分布及果实数量与植株年龄和生境的关系四合木的果实常单生于叶腋,具有4个不开裂的分果瓣。分果瓣呈新月形,长4~8 mm,宽2~4 mm。种子呈镰刀状披针形,灰黄色至褐色,长2~3 mm,宽1.5 mm。种皮革质,表面密被褐色颗粒。种子细小,无胚乳。
据调查区域内大量野外调查发现,3 a生以内四合木植株均未开花结实,据此,以3 a为一龄级,对所调查的107株(丘地60株,滩地47株)四合木进行龄级划分(如表 2),分别统计各龄级四合木平均果实的数量,绘制果实数量与龄级相关关系图(图 1)。
|
|


|
图 1 四合木果实数量随龄级变化 Fig. 1 The changes of fruit number with age class |
图 1表明,无论丘地、滩地,四合木种群平均每株果实的数量随年龄的变化规律基本一致,都是中间高,两头低,随年龄的变化曲线呈山峰型。具体来说,丘地四合木在2龄级时就有少部分植株开花结实,初始结实年龄为4 a生;而滩地四合木初始开花结实年龄在2龄级之后。共同表现是:在2~6龄级范围内,随着龄级增大,四合木的结实量增加,6龄级以后,随着龄级的增大,结实量逐渐减少。结实高峰期在5~9龄级。
四合木单株结实量在树冠的不同部位分配上存在一定的差异。为了揭示四合木结实量在树冠空间分布的规律,对在丘地调查的55株3~11龄级的四合木和在滩地调查的47株同一龄级的四合木果实数量,区分树冠的南侧、北侧、上、中、下等不同部位分别进行了统计,结果见表 3、4。
|
|

|
|
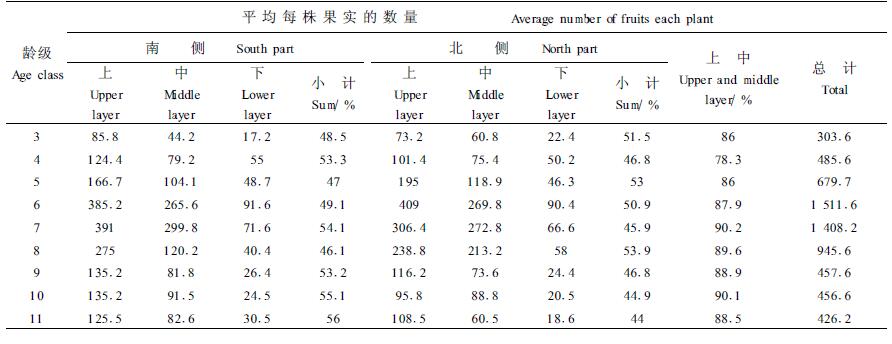
从表 3、4可以看出,丘地、滩地四合木种群各龄级平均单株果实数量分布无显著差异,丘地和滩地四合木冠层南侧与北侧的果实数量分布也无显著差异。在调查区域四合木株高一般不超过70 cm, 在草原化荒漠地区多建群种、优势种或亚优势种,所在群落盖度一般小于0.5,群落内南北向光照皆较充足。
按垂直分,丘地各龄级四合木冠层中、上部占全株果实数量的77.5%~92.4%,滩地各龄级四合木冠层中、上部占全株果实数量的78.3%~90.2%,上、中部显著高于下部。产生这种现象的原因主要取决于光照条件和花的分布,四合木的花常单生叶腋,虽然群落内光照充足,但每个植株冠上部对中下部具有一定遮荫作用。充足的光照条件有利于花芽的分化,所以就群体结实水平而言,出现了树冠上部>中部>下部,上、中部高于下部这种现象。
2.2 果实重量与植株年龄和生境的关系种子的质量与种子的活力有关。活力旺盛的种子也就是无病的、能够萌发的饱满种子,它们对环境条件的要求相对较低,一般情况下都能出土萌发成幼苗,成为种群更新的补充库。因此,种子质量的好坏是影响种群未来命运的关键因子。了解种子质量和活力的途径主要有物理性状(种子重量和大小)和生理指标。通常,大粒种子或重量大的种子比小粒种子具有较充实的贮藏物质(傅家瑞,1985)。就四合木而言,四合木种子细小,其千粒重平均为1.112 1 g(吴素琴等,1994)。并且由于种子中有一定比例的空粒,为此选用重量作为评价四合木质量的指标。
将丘地64株及滩地65株四合木果实干重分别统计,以龄级为横坐标,各龄级平均每株果实的干重为纵坐标绘制果实干重与龄级关系曲线,如图 2。图 2所示,四合木果实的重量在不同龄级的植株间差别很大,各龄级平均单株果实重量随龄级呈单峰型变化。在4~6龄级以内,各龄级平均单株果实干重基本上随龄级增加而增加,丘地在6~7龄级平均单株果实重量处于高峰期,滩地5~8龄级平均单株果实重量处于高峰期,7龄级以后,果实干重随龄级增加而下降,这与果实数量随龄级的分布趋势基本一致。四合木种子重量轻、饱满种子数量少,是一种对恶劣环境的适应对策,这样不仅可以节约能量,保证植株的生长需求,而且有利于在种间竞争中处于优势。

|
图 2 丘地、滩地各龄级平均每株果实干重 Fig. 2 The average dry weight of fruits each plant in different age class of T. mongolica in the plots of hill and beach |
经观察,四合木为两性花,单生于叶腋或1~2朵生于短枝上。前人记载萼片4,花瓣4,雄蕊8,分2轮排列,心皮4。观察发现有以下变异:萼片4~5,花瓣4~5,雄蕊8~10,心皮4~5。
四合木的花期为5月上旬—6月初,单朵花的花期约为4~5 d,整株开花可持续8~15 d。一般上午花盛开,下午多数萼片、花瓣呈半合拢状;花将开时,花丝和花柱就已伸长,此时柱头高出花药2~3 mm;盛开时,花丝进一步伸长,与柱头近等长,内轮4个雄蕊花药先开裂,开裂方式为背向纵裂,并紧围靠着雌蕊柱头,柱头有分泌物,可与花药粘在一起,外轮4个雄蕊花药至少晚1 d开裂;花将谢时,花药全部开裂,花粉散毕之后,萼片、花瓣又闭上,过几天,萼片、花瓣即脱落。
2.4 花果转移率及植株年龄和生境的关系在丘地和滩地分别选取45株四合木标准植株,调查植株花蕾数和初果数,并计算花果转移率,统计结果见表 5。
|
|

由表 5所示,滩地四合木单株花果转移率随龄级变化皆高于丘地,6龄级以后,随着年龄增大,花果转移率下降。无论丘地还是滩地,四合木平均每株花果转移率随龄级变化具有波动,有2个小的高峰期。丘地2个高峰期为2龄级和5龄级,滩地2个高峰期为3龄级和5龄级。从所调查丘地、滩地各45株四合木花果转移率看,在自然状态下,丘地四合木植株花果转移率在32.98%~68.33%之间,平均转移率为57.36%, 各龄级平均花果转移率在54.39%~61.06%之间;滩地四合木植株花果转移率在40.82%~83.29%之间,平均为68.76%, 各龄级平均花果转移率在64.49%~71.43%之间。
在相同龄级,不同生境、不同群落类型中的四合木种群的花果转移率存在差异。杭锦旗滩地四合木种群在27 a生以内各龄级的花果转移率比相同龄级丘地的花果转移率高。据徐庆等(2001)研究,滩地四合木种群在27 a生内各龄级的生殖值比相同龄级的丘地生殖值高。在生境条件较差的群落中,四合木种群生殖值、花果转移率高,这是其适应恶劣环境所采取的生殖对策。
2.5 结籽率与植株年龄和生境条件的关系将所调查的丘地的55株及滩地的40株四合木单株的结籽率分别统计,以龄级为横坐标,各龄级平均单株结籽率为纵坐标,绘制结籽率随年龄变化图(图 3)。

|
图 3 丘地、滩地各龄级平均单株结籽率 Fig. 3 Average seed bearing percentage each plant in different age class of T. mongolica in the plots of hill and beach |
图 3表明,在自然条件下,不论丘地还是滩地的四合木种群各龄级单株的结籽率较低。丘地各龄级平均单株结籽率为1.40%~2.80%,滩地各龄级平均单株结籽率为1.26%~2.65%,除3龄级(7~9 a生)外,滩地各龄级的结籽率皆低于丘地的结籽率。四合木单株结籽率随龄级变化呈双峰型。在4~6龄级,随年龄增加,结籽率略有增高,6龄级以后,随着年龄增大,结籽率下降。据吴树彪(1990)研究,四合木开花后各期均有胚珠败育。刘果厚等(1993)对40株生长发育良好的四合木,1990年4月—1991年7月进行了定点定株施肥和浇水,结果表明,施肥和浇水不但可以使四合木的结实量和千粒重提高,其生长量也有所提高。说明结籽率低与分布区的土壤贫瘠和干旱缺水有密切的关系。丘地土壤含水量略高于滩地,丘地四合木的结籽率也略高于滩地,进一步证明结籽率高低与土壤含水量有关。
为了探讨四合木结籽率低的原因,1998年5月,在四合木分布区杭锦旗,进行了套网、套袋、去雄试验。具体做法是:在四合木开花前(花蕾期),随机选取8株四合木个体,把整个花枝套上尼龙网(网眼大小为1 mm×1 mm, 完全隔离了昆虫传粉)进行套网试验,结果如表 6。
|
|

从表 6看出,四合木种群为虫媒传粉。野外定点观察发现,四合木的传粉昆虫有蚁类和蜂类,传粉昆虫的报酬是花盘分泌的蜜液。蚂蚁的活动范围小,一般只在同一枝条的花间访问,在一朵花上停留约2~3 min,且访花频率与植株下有无蚁穴有关,有的植株下无蚁穴,则看不到蚂蚁传粉;蜂类的数量极少,而且只在无风晴朗的天气时才出现,在一朵花上停留的时间不长,几秒到十几秒,在一丛间停留数分钟,然后突然飞走。一般下午3:00后,就看不到蜂类访花。另外,这些蜂类同时也是与四合木同花期的阿氏旋花的传粉者。因此,认为四合木结实率低的主要原因是传粉过程受到障碍,即传粉媒介(昆虫)的访花频率极低,使得柱头上的花粉量受到限制。这是结籽率低的内因。
3 结论杭锦旗丘地四合木种群初始开花结实的年龄约为4 a生,四合木种群的果实数量、果实重量从低龄组到高龄组,呈现出一种低→高→低的趋势。在4~18 a生范围内,随着年龄增大,结实量增加;到18 a生以后,随着年龄增加,结实量逐渐减少;13~27 a生为结实高峰期。
就群体结实水平而言,四合木种群结实量在树冠上的空间分布为:树冠上部>中部>下部,树冠中、上部显著高于下部。南北侧差别不大。这与树冠中、上部光照优于下部,南北方向光照皆较充足有关。
在自然状态下,四合木单株花果平均转移率为57.36%~68.76%, 结籽率为1.26% ~2.80%。结籽率低的主要原因是:1)传粉过程受到障碍,即传粉媒介(昆虫)的访花频率极低,使得柱头上的花粉量受到限制。这是结籽率低的内因。2)土壤水分供应不足,从而使胚珠不能正常发育。这是导致结籽率低的外因。
曹坤方. 1993. 植物生殖生态学透视. 植物学通报, 10(2): 15-23. |
方炎明. 1996. 植物生殖生态学. 济南: 山东大学出版社.
|
傅家瑞. 1985. 种子生理. 北京: 科学出版社, 76-86.
|
刘果厚, 周世权. 1993. 内蒙古特有植物四合木生物学特性及濒危原因的研究. 内蒙古林学院学报, 15(1): 44-49. |
苏智先, 张素兰, 钟章成. 1998. 植物生殖生态学研究进展. 生物学杂志, 17(1): 39-46. |
王迎春, 侯艳伟, 张颖娟, 等. 2001. 四合木生殖对策的研究. 植物生态学报, 25(6): 699-703. DOI:10.3321/j.issn:1005-264X.2001.06.009 |
吴树彪, 屠骊珠. 1990. 四合木胚胎学研究. 内蒙古大学学报, 21(2): 277-283. |
吴素琴, 李克昌, 杨瑞全. 1994. 四合木种子特性的测定研究. 草业科学, 11(3): 29-31. |
谢宗强, 陈伟烈, 胡东, 等. 1998. 濒危植物银杉的结实特性及动物对果实的危害性. 植物生态学报, 22(4): 319-326. DOI:10.3321/j.issn:1005-264X.1998.04.005 |
徐庆, 刘世荣, 臧润国, 等. 2001. 中国特有植物四合木种群的生殖生态特征:种群生殖值及生殖分配研究. 林业科学, 37(2): 36-41. |
徐庆, 臧润国, 刘世荣, 等. 2000. 中国特有植物四合木种群结构及动态研究. 林业科学研究, 13(5): 485-492. DOI:10.3321/j.issn:1001-1498.2000.05.005 |
朱宗元, 雍世鹏, 刘钟龄.四合木荒漠(Form. Tetraena mongolica).见: 中国科学院内蒙古宁夏综合考察队编.内蒙古植被.北京: 科学出版社, 1985: 687-690
|
Abrahamson W G. 1982. On the comparative allocation of biomass, energy, and nutrients in plant. Ecology, 63(4): 982-991. DOI:10.2307/1937238 |
Aker G L. 1982. Regulation of flower, fruit, and seed production by a monocarpic perennial, Yucca whipplei. J Eco, 7: 357-372. |
Bazzaz F A. 1979. Contribution to reproductive effort by photosynthesis of flowers and fruits. Nature, 279: 554-555. DOI:10.1038/279554a0 |
Douglus D A. 1981. The balance between vegetative and sexual reproduction of Mimulus primuloides(Scrophulariceae) at different altitudes in California. J Ecol, 69: 295-310. DOI:10.2307/2259832 |
Hegazy A K. 1992. Age-specific survival, mortality and reproduction, and prospects for conservation of Limonium delicalumlum. J App Ecol, 29: 549-557. DOI:10.2307/2404462 |
Harper J L. 1997. Population biology of plants. London: Academic Press, 1-200.
|
Silvertown J W. 1982. Introduction to plant population ecology. London and New York: Longman.
|
Silvertown J W, Dous J L. 1982. Introduction to plant population ecology. Oxford: Blackwell Scientific Publication.
|
Steven D D. 1991. Experiments on mechanisms of establishment in old-field succession:seedling emergence. Ecol, 72: 1066-1075. DOI:10.2307/1940606 |
Willson M F. 1983. Plant reproductive ecology. New York: John Wiley and Sons.
|