 2003, Vol. 39
2003, Vol. 39文章信息
- 臧润国, 杨彦承, 林瑞昌, 杨秀森.
- Zang Runguo, Yang Yancheng, Lin Ruichang, Yang Xiusen.
- 海南霸王岭热带山地雨林森林循环与群落特征研究
- STUDY ON THE FOREST CYCLE AND COMMUNITY CHARACTERISTICS IN A TROPICAL MONTANE RAIN FOREST IN BAWANGLING, HAINAN PROVINCE
- 林业科学, 2003, 39(5): 1-9.
- Scientia Silvae Sinicae, 2003, 39(5): 1-9.
-
文章历史
- 收稿日期:2001-11-01
-
作者相关文章


2. 中国林业科学研究院资源信息研究所 北京 100091;
3. 海南省霸王岭林业局 昌江 572722
2. Institute of Forest Resources and Information Technology, CAF Beijing 100091;
3. Bawangling Forestry Bureau of Hainan Province Changjiang 572722
自英国著名生态学家Watt(1947)开创性的工作后,人们逐渐认识到植物群落是由不同性质的斑块所构成的镶嵌复合体, 自然干扰在森林的结构、动态和多样性维持中具有非常重要的作用(Pickett et al., 1985; Denslow, 1987; Everham et al., 1996; Moloney et al., 1996; Valverde et al., 1997)。受到Watt学术思想的影响,Whitmore进一步在热带雨林的研究中拓展了植物群落内斑块动态的思想,并发展为森林循环(forest cycle)的理论(Whitmore,1989;臧润国等,1999)。这一理论认为,任何森林中都存在着由干扰所驱动的林分斑块的循环变化过程,我们可以大致将其划分为林隙阶段(gap phase)、建立阶段(building phase)、成熟阶段(mature phase)和衰退阶段(degenerate phase)(Whitmore, 1988;1989)。森林中一定时期内某个地段是以林隙阶段为主,而另外的地段是以建立阶段为主,还有的地段则可能是以成熟阶段或衰退阶段为主。随着时间的推移,林隙阶段的斑块就会逐渐发育为建立阶段,建立阶段的斑块会发育为成熟阶段,成熟阶段会发育为衰退阶段,衰退阶段的斑块则在外界干扰因子(风、火、病虫害、树木衰老死亡等)的触发下,又会重新转化为林隙阶段。因此,在整个森林景观中,不同的斑块通过干扰因子的作用,就形成了此起彼伏的动态变化过程。随着斑块的动态变化过程,不同斑块内的生态因子及其组合不同,在其中更新、生长和繁衍的植物的种类、数量、大小等就会不同,因此,随着森林循环过程的进行,森林内树木的种群特征和斑块的群落特征就会随之发生相应的动态变化。斑块动态的思想引起了生态学范式的变化,对当今生态学理论产生了重要的影响(邬建国, 2000)。森林循环的理论,为我们研究群落内斑块的动态规律提供了新的思路。国外有关森林循环与群落动态规律方面的研究还不多,Whitmore(1978;1989a;1989b)研究过东南亚和Solomon群岛热带雨林的森林循环及群落动态,Martínez-Ramos (1988)对墨西哥的热带雨林也做过有关森林循环斑块阶段的划分,但与森林循环有关的群落动态过程还涉及很少。我国在森林循环的研究方面,还刚刚起步,臧润国等(1999)对红松林的斑块动态做过很初步的分析,但有关森林群落特征随森林循环过程的变化是一个目前尚未有人涉足的空白领域。本文以我国海南省典型的热带山地雨林为研究对象,在分析山地雨林一般群落特征的基础上,以森林循环理论为指导,探讨种群或群落特征与森林循环的关系,进而为阐明自然干扰与森林群落内的斑块镶嵌格局和森林生物多样性的形成与维持的关系奠定基础。
1 研究地与研究方法 1.1 研究地点本研究的调查地点设在海南省霸王岭国家级自然保护区的原始热带山地雨林内,其地理坐标为北纬18°50′~19°05′,东经109°05′~109°25′。霸王岭国家级自然保护区,位于海南省昌江县与白沙县的交界处,面积约72000 hm2,其中1980年建立的长臂猿(Hylobates concorloris)保护区位于800 m以上的山地,覆盖面积约2500 hm2,是海南省保护最好且最为典型的热带山地雨林。我们具体的调查地点在保护站旧址的东北侧,坡度大部分在5°~10°左右。土壤为山地黄壤。气候为热带季风气候, 年平均温度23.6℃,年均降水量1500~2000 mm。调查地点主要是以陆均松(Dacrydium pierrei)、线枝蒲桃(Syzygium araiocladum)等为优势的热带山地雨林群落。
1.2 研究方法野外调查采用格局分析的方法。在霸王岭自然保护区保护站旧址东北侧的固定样地内(余世孝等,1993),选择东西长100 m,南北长100 m的调查样地,在这1 hm2的样地内,密布5 m × 5 m的相邻网格小样方,共400个。从样地西北角的第一个小样方开始,依次对每个小样方进行调查。调查前,首先判定其在森林循环中所处的阶段。然后对每个小样方内高度在1.5 m以上的所有乔、灌木进行每木调查,即辨认样方内每棵树木的种名,测定其胸径和树高。在野外判定各样方所属森林循环斑块阶段的标准如下:(1)林隙阶段(G):当在林隙中时可明显看出其内的光亮度较大,林隙内一般可看到以掘根、折干或枯立等形式出现的形成木(gap maker)或腐朽树桩的痕迹(臧润国等,1999)。林隙内绝大部分树木的高度都达不到林隙周围主林层树木高度的1/2,林隙内树木的冠层也没有与周围主林层的树冠相接触,一般没有或很少有高度在8 m以上或胸径在10 cm以上的树木。(2)建立阶段(B):是由林隙阶段发育而来,一般已看不到林隙形成木及其痕迹,林分内的光亮度已远不如林隙内大,斑块内树木冠层的高度这时已达到与周围主林层树冠中下部相接触的地步,但如站在林分内,仍能看出其和周围大树的高度有一些差别,即在此阶段林分的上层仍可看出其周围主林层围绕的空隙轮廓。这个空隙与林隙阶段的相比,已被大大抬到了高处,深度也浅了许多。斑块内有不少高度在8 m以上或胸径在10 cm以上的树木,但高度在14 m以上或胸径在20 cm以上的树木还不多。(3)成熟阶段(M):是建立阶段林分进一步发育的结果,此阶段林分的上层树木已有很多处于主林层,这时林分的冠层即为主林层。斑块内有较多高大树木,基本上没有明显衰老的树木。高度在14 m以上或胸径在20 cm以上的树木已有很多,但很少有胸径在50 cm以上的树木。(4)衰退阶段(D):是成熟阶段林分逐渐走向死亡的过渡阶段,这个阶段的树木有许多是又高又粗的大树,且有不少粗大树木的高度越过了主林层,即已成为了所谓的超冠层树木(emergent trees),但这个阶段斑块的明显特征是大树的活力已大为减少,有许多大树都表现出叶子稀疏、折枝断梢、秃顶和树干上有明显的腐坏斑片或树洞、树干空朽等衰老的特征,处于这个阶段的大部分斑块中都有1株或几株胸径在50 cm以上的大树。调查中当一个样方同时跨越上述两个阶段以上的斑块时,看哪个发育阶段的斑块在样方中所占的比例最大,就将其归入哪个发育阶段的斑块类型。这样的网格样方调查方法,有似微积分的方法,即首先将1 hm2的森林“微分”为400个小网格,然后判定每个小网格内所属的发育阶段斑块类型。当完成对每个小样方的调查后,我们就可将同类型斑块的样方合并在一起,比较林隙(G)、建立(B)、成熟(M)和衰退(D)各阶段斑块内或四个阶段合在一起的种群与群落特征了。这个过程,可称其为“积分”的过程。
1.3 计算方法对于全林分而言,以所调查的1 hm2样地内每个5 m×5 m的小样方为基础, 统计每个树种出现0, 1, 2, 3, ……个个体的样方数, 并以此为基础, 计算它们的几个聚集强度指标, 其中K=m 2 /(v-m)为负二项系数, K值愈小, 聚集强度越大;C=v/m为扩散系数, C=1时种群为随机分布, C>1为聚集分布;CA=1/K为Cassie指标, CA=0时为随机分布, CA>0为聚集分布, CA < 0为均匀分布;I=(v/m)-1为丛生指标, I=0时为随机分布, I < 0为均匀分布, I>0时为聚集分布。上述公式中,m为样本的平均值,v为其方差。
我们可把霸王岭热带山地雨林中调查到的400个样方中的每一个样方, 看作是一个不同的资源状态, 即在热带山地雨林的水平异质性基质上, 每个样方内的资源状态及其对每个树种的有效性是不同的。根据400个5 m × 5 m小样方的调查数据, 以每个树种在每个小样方内出现的个体数为基础, 应用Levins提出的Simpson指数计算公式
森林循环由林隙阶段经建立阶段到成熟阶段最后达到衰退阶段, 伴随着这种斑块发育阶段的动态变化, 不同阶段斑块内的物理环境和生物环境也必然会发生相应的变化, 因此, 对于不同的树木种群来说, 森林循环的每一个阶段就可以被看成是一个相对不同的资源状态, 即G、B、M、D阶段分别是对树木种群可利用或占据性不同的四种不同的资源状态,四个阶段可以当作是树木种群在热带山地雨林群落中水平空间的资源利用谱。按照生态位的定义,不同树木种群通过资源利用,就形成了它们在森林群落中的生态位关系。应用Levins计算式, 并分别以不同斑块内每个树种出现在四个不同阶段斑块内的个体数或胸高断面积为基础计算出热带山地雨林中主要树种的水平生态位宽度,此时公式中S为森林循环的阶段数。
热带山地雨林森林循环不同阶段斑块之间的相似性群落系数运用Sresen公式计算, 
从计算出的霸王岭热带山地雨林内不同树种的水平生态位宽度值的大小可以看出, 不同树种对山地雨林内水平空间上的不同小斑块的利用或占据能力不同, 故而表现出其水平生态位宽度的巨大分化, 依据生态位宽度值的大小范围, 可将热带山地雨林内的树种分为以下几组:①生态位宽度值在0.7以上的树种, 可称其为是对山地雨林水平斑块生态资源利用很充分的广布种, 即它们在山地雨林的每个斑块内大都有分布, 且个体数在不同小斑块内的分布均匀。其中有不少是热带山地雨林内第二、三层的中小乔木树种如线枝蒲桃等, 也有不少是热带山地雨林中常见的灌木树种如九节(Psychotria rubra)、柏拉木(Blastus cochinchinensis)等。这一类的树种在所调查的热带山地雨林中共有17种;②生态位宽度在0.5~0.7之间的树种, 可称其为对热带山地雨林水平生态资源利用较充分的树种。在调查的树种中共有30种, 它们对山地雨林不同斑块内的生态资源有较充分的利用, 但不如第一类树种, 如油丹(Alseodaphne hainansis)、陆均松等;③生态位宽度在0.3~0.5之间的树种, 可称其为对热带山地雨林水平生态资源利用不充分的树种。共有29种, 它们只对热带山地雨林的水平生态资源有一定程度的利用, 但不充分,如侯氏新木姜(Neolitsea howii)、锈毛杜英(Elaeocarpus howii)等;④生态位宽度在0.1~0.3之间的树种, 可称其为对热带山地雨林水平生态资源利用极不充分的树种, 共有29种。它们中的每个种,大都只在热带山地雨林的少部分斑块内分布,且个体数也很少,大部分的水平斑块资源对它们来说是不可利用或没有利用,如子凌蒲桃(Syzygium championii)、羽叶泡花(Meliosma angustifolia)等;⑤生态位宽度在0.1以下的树种, 是热带山地雨林中的低密度稀有种, 如凸脉冬青(Ilex kobuskiana)等共有30种。它们在热带山地雨林中仅有零星分布,每当它们出现时,也都是以极少数的个体出现,因此,这类树种与群落中的其它树种相比,是偶见种或稀有种,它们仅零星出现在少数类型的水平斑块内,呈随机分布状态。
2.1.2 垂直生态位宽度根据不同树种垂直生态位宽度的大小可将热带山地雨林中的树种划分为以下几类:①垂直生态位宽度值在0.7以上的树种, 是那些对山地雨林垂直空间有很充分利用的种, 它们的个体数较多且在垂直空间分布较均匀, 可称其为对山地雨林垂直空间利用很充分的树种, 如油丹、黄叶树(Xanthophyllum hainanense)等9种;②垂直生态位宽度值在0.5~0.7之间的树种, 可称其为对山地雨林垂直空间利用较充分的树种, 如陆均松等38种;③垂直生态位宽度值在0.3~0.5的树种, 可称为是对热带山地雨林垂直空间资源利用较不充分的树种, 如华润楠(Machilus chinensis)、柏拉木等43种;④垂直生态位宽度值在0.3以下的树种, 可称为是对热带山地雨林垂直空间资源利用很少的树种, 如粗叶木(Lasianthus chinensis)等47种。
2.2 主要树种的空间分布格局分析 2.2.1 主要树种在森林循环不同阶段斑块内出现的频度通过对海南霸王岭热带山地雨林主要树种在森林循环不同阶段斑块内出现频度的统计表明,不同树种在森林循环的不同阶段出现的频度有较大的差异,有些树种只在森林循环的某一阶段出现,在其它阶段不出现,如拟赤杨只出现在G阶段, 在其它阶段则没有出现。这可能是由于这些树种的耐荫性很小,只能出现在有较多光照的树隙内。有些树种则只出现在非林隙阶段,即除林隙外,它们在森林循环的其它阶段都有出现,这些树种大多是较为耐荫的非林隙树种,如肖柃(Cleyera obscurinervis)。还有些树种在森林循环的各个阶段都出现,且出现的频度都较大,如线枝蒲桃、九节等,它们是生态适应幅度较宽广的树种。在森林循环的不同阶段,各个树种出现的频度不同,而就同一树种而言,它们在森林循环不同阶段斑块内出现的频度大小也有很大的变化,表 1明显的显示了这一点。
|
|

不同树种在森林循环不同阶段斑块内出现的频度不同,从而表现出其水平格局的差异。以400个小样方为基础,计算出热带山地雨林内树种的聚集强度指数(如表 2所示),可以看出, 霸王岭热带山地雨林中不同树种表现出不同的空间分布类型。在所调查的1 hm2热带山地雨林中, 共调查到乔、灌木树种138种, 其中呈随机分布的有27种, 均匀分布的有50种, 聚集分布的有61种。逐个分析后可以看出,占优势的乔灌木树种如陆均松、线枝蒲桃等大都呈聚集分布的格局,个体数量中等的树种大都呈均匀分布的格局,而那些个体数量少的低密度种则大都呈随机分布的格局。呈聚集分布树种的密度较高,而均匀或随机分布种的密度则相对较低。
|
|

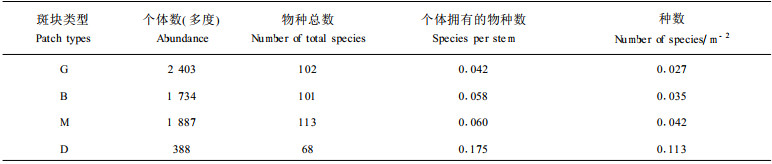
表 3给出了霸王岭热带山地雨林森林循环不同阶段斑块的物种数、个体数、密度的一些特征。可以看出, 处于森林循环不同阶段斑块内的物种数及其个体数特征不同, 从四个阶段的物种总数上来看, G和B阶段的几乎一样, M阶段的最高, 而D阶段则大为降低, 几乎降为M阶段的种数的一半。从个体数的总数,即多度来讲,处于G阶段的个体数多度最大, D阶段的最小, 而B和M阶段的较接近。但这种不同阶段斑块的比较,没有考虑面积的作用,因为各阶段斑块在森林景观中的面积相差很大,所以这种比较的可比性不强。为比较不同斑块内物种数的多少,国外有些学者(Denslow, 1995)分别采用了平均每个个体所拥有的物种数, 即用物种数除以调查到的对应的个体数(species per stem), 以及单位面积的物种数(species per area)。从表 3可以看出, 在考虑了密度和面积效应后, 不同斑块之间的物种密度, 无论是以平均每个个体拥有的物种数, 还是以每平方米面积上拥有的物种数来说, 海南霸王岭热带山地雨林的物种密度都随森林循环阶段的增加, 而呈现逐渐增加的趋势, 特别是由M到D阶段时, 种密度增加了近3倍。
|
|
分别统计霸王岭热带山地雨林森林循环不同阶段斑块内的拥有1, 2, 3, …, 14, ≥15个个体的物种数, 并绘制种—个体数曲线, 如图 1所示。可以看出, 随着个体数的增加, 相对应的树种数也在逐渐减少, 尤其是由单个体和双个体所对应的物种数较多, 3个以上个体对应的树种数则呈现出明显的下降。个体数≥15以上的树种数是个体数在15及其以上的所有树种数的累加,并不代表对应于某一个体数的物种数,所以其物种数也有一定的数量。

|
图 1 霸王岭热带山地雨林的种-个体数曲线 Fig. 1 The species-individuals curves of different patch types of forest cycle in the tropical montane rain forest |
不同阶段斑块之间的相似系数如表 4所示。可以看出, 不同阶段斑块之间在物种组成上的相似性程度不同, 其中任何两个相邻阶段斑块之间的相似性都高于不相邻斑块间, D阶段斑块与其它三类斑块之间的相似性最小。
|
|

以森林循环的不同阶段斑块为资源单位,分别以个体数和胸高断面积为基础,计算出霸王岭热带山地雨林不同树种的生态位宽度,如表 5所示。从表 5可以看出, 不同树种的生态位宽度有很大的变异, 那些在森林循环四个阶段斑块内都有出现, 且出现的个体数或胸高断面积在不同斑块间分布相对均衡的树种如线枝蒲桃、九节、黄叶树等的生态位就较宽,而只在某些阶段斑块出现或虽在每个斑块都出现,但个体数或断面积在不同斑块间分布数量不均衡的物种如陆均松(只出现在M、D阶段)等,其生态位就相对小,而那些个体的数量很少,只在林分中呈随遇分布的种,如肖柃、侯氏新木姜等,它们大都是热带山地雨林中的低密度种群,其生态位宽度一般都很小。不同树种的生态位宽度不仅与不同阶段斑块内的生态因子有关,而且也与树木本身的生物学生态学特性有关。根据以个体为基础计算的生态位宽度数值的大小变化,可将霸王岭热带山地雨林中的树种分为以下几类:①对森林循环各阶段生态资源利用充分的广生态幅树种:其生态位宽度B值在0.7以上, 包括线枝蒲桃、三角瓣花(Prismatomeris tetrandra)等30种。②对森林循环各阶段生态资源利用较充分的树种,其生态位宽度在0.5~0.7之间, 包括黄叶树、盘壳栎等35种。③对森林循环各阶段生态资源利用不充分的树种,其生态位宽度在0.2~0.5之间,包括锈毛杜英、陆均松等26种。④对森林循环不同阶段生态资源利用极不充分的树种,它们大都是只在森林循环的某个阶段以极少数的个体存在的低密度种,其生态位宽度在0.2以下, 包括肖柃、拟赤杨(Alniphyllum fortunei)等27种。在我们所调查的热带山地雨林中有100多个树种, 由于这些树种各自的形态及生态特性不同, 从而表现出其个体的大小及其在森林群落中的地位不同。由于不同树种之间个体大小差异很大,完全用个体数为基础来计算不同树种的生态位大小,有很大程度的不可比性,但目前在森林群落的研究中,还没有更好的办法来解决这一问题。不同树种胸高断面积的大小在一定程度上能表明其对环境资源的利用能力的大小,树木的胸高断面积越大,在一定程度上表明其对环境资源的利用能力越大,在群落中的优势度也就越大。为此,我们又以森林循环不同阶段的斑块为基本资源单位,用树木的胸高断面积为基础计算了霸王岭热带山地雨林中不同树种的生态位宽度列于表 5中。从表 5可以看出,一般用断面积为基础计算的生态位宽度要小于以个体数为基础计算的生态位宽度。图 2和图 3分别是热带山地雨林的树种数在以个体数和胸高断面积为基础的生态位宽度上的分布情况, 可以看出, 如果以个体数为基础, 则生态位宽度在0.6~0.8和0.2以下的树种最多, 而如果以断面积为基础, 则树种数随生态位宽度的增加而呈现逐渐下降的趋势, 在0.8以上没有, 而在0.2以下的树种则占了一半多。
|
|


|
图 2 树种数在以个体数为基础计算的生态位宽度上的分布 Fig. 2 Relationship between species richness and niche breadth(calculated by individuals) |

|
图 3 树种数在以胸高面积为基础计算的生态位宽度上的分布 Fig. 3 Relationship between richness and niche breadth(calculated by basal areas) |
如果按生活型或个体的大小来分析的话,可以看出霸王岭热带山地雨林中绝大部分的灌木树种水平生态位宽度较大,其次是第2、3层的中、小乔木树种,大乔木树种,它们在群落中的重要值或优势度虽然很大,但它们的水平生态位宽度居中,而那些低密度、稀有种的水平生态位宽度最小。水平生态位宽度的这种变化可能与不同树种的水平分布格局有很大的关系,如灌木和中、小乔木在群落中的大多数斑块中都有出现,分布相对均匀,对各类生境都有较大的利用,故其生态位宽度较大,而大乔木树种则由于其个体较大,分布的均匀性较小,在某些斑块中就较少出现,故其生态位宽度居中。而那些低密度稀有种,只局限在某些特定类型的生境小斑块中,故其水平生态位极其狭小。从垂直生态位宽度的大小我们可以看出,绝大部分的灌木或小乔木树种大都对垂直生态资源的利用不太充分,而在群落中占优势的乔木大都对垂直生态资源的利用较充分。可以看出,热带山地雨林中不同生活型的树种在利用垂直和水平生态资源上是相对分离的,绝大部分的灌木或小乔木树种对水平生态资源利用较为充分,对垂直生态位则利用较不充分,而中、大乔木则正好相反。不同生活型的树种,通过对不同空间中生态资源的利用程度的不同而在群落中具有独特的生态位,不同树种生态位的相对分离是它们能够长期共存的基础,也是热带山地雨林物种的多样性得以维持的基础。
森林循环不同阶段的斑块为不同特性的树种提供了不同的生长发育条件。不同树种在森林循环各阶段斑块内出现频度的高低差异,反映了不同阶段斑块内生态环境的不同,同时也表明了不同特性的树种对森林循环不同阶段斑块生境条件的选择和适应特性不同。
如果分别将种数/个体数和种数/面积比值倒过来,我们就可以看出,随着森林循环过程的推进,热带山地雨林中每个树种所拥有的个体数是逐渐减少的,且每个树种所拥有的空间面积也在不断减少。这可能是由于随着森林循环进程的推进,森林斑块内不同种群之间以及同一种群内的不同个体之间的竞争在不断加大,到一定阶段后,只有一部分树种的少部分个体进入到成熟和衰退阶段(臧润国等,2001),且到了这些阶段的个体体积也比以前的几个阶段有非常大的增加,这样每个个体拥有的空间较大,而平均每个树种所拥有的个体数以及面积就会大为减少了。
逐个分析各物种在不同斑块内分布的个体数时发现,个体数≥15以上的树种在G、B、M、D各阶段分别有27, 25, 27和5种, 其中相应的灌木分别有10, 8, 8和4种, 第二、三层的乔木树种线枝蒲桃等也拥有较多的个体, 而山地雨林中绝大多数的优势树种如陆均松、黄叶树等在各类斑块内拥有的个体数大都有很大的变化。我们在1 hm2的热带山地雨林中, 调查到乔灌木树种138种, 其中各类斑块中都有大量的低密度树种, 1~3个个体的物种数在G、B、M、D各斑块中的树种数分别为37、37、50和44,分别占各斑块内树种总数的36%、37%、44%和65%。不同斑块内的种-个体数曲线类型基本相似,但G和B阶段的单个体和双个体的种类相对较少, 而D阶段多个体(≥15)的物种数则明显少于其它三个阶段。
森林循环不同阶段斑块之间组成相似性的大小在一定程度上表明了热带山地雨林森林循环斑块动态的连续性,也就是说随着森林循环过程的推进,斑块内的物种组成是逐渐的,而不是间断式或跳跃式变化的。这在一定程度上可通过相邻阶段斑块之间的相似性程度表明出来。相邻斑块之间共有种的数量一般都大于不相邻斑块之间共有种的数量。
出现图 2,图 3的结果的原因可能是, 热带山地雨林中大部分树种都为矮小或低密度的树种(臧润国等,2001), 故其胸高断面积大都很小, 且在不同斑块之间的数量分布不均匀, 故其以断面积为基础的生态位宽度就很小。另外,有些树种如陆均松等,它们的胸高断面积和个体数虽然都很大,但由于其只集中分布在森林循环的后两个阶段的斑块内,断面积在不同斑块类型之间的分布极不均匀,故其生态位宽度也很小。那些在森林循环的不同阶段都出现,且个体数或断面积在不同类型斑块之间分布均匀的树种,如线枝蒲桃等,其无论以个体数,还是以胸高断面积为基础的生态位宽度都是较大的,它们对森林循环不同阶段斑块类型内的资源都有较充分的利用,其生态适应幅在热带山地雨林中是较宽的。
本研究的结果表明,热带山地雨林群落是由不同性质的林分斑块所组成的镶嵌复合体,森林植物种群或群落特征在不同时空上的差异(Moloney et al., 1996),表明了森林群落的异质性特点(Lertzman et al., 1996)。森林循环是群落内异质性产生的重要过程,是不同生活史特性树种在群落中实现生态位分化和长期共存的重要机制,也是森林生物多样性和生态系统的动态稳定性得以维持的基础。实际上群落内组织结构的小尺度差异正在引起生态学家的注意(叶万辉等,1998;Van der Maarel et al., 1995)。
彭少麟. 1996. 南亚热带森林群落动态学. 北京: 科学出版社, 332-373.
|
邬建国. 2000. 景观生态学-格局、过程、尺度、等级. 北京: 高等教育出版社.
|
叶万辉, 黄忠良, 孔国辉, 等. 1998. 鼎湖山南亚热带常绿阔叶林定位研究(Ⅳ)锥栗、黄果厚壳桂群落组织结构的小尺度差异. 北京: 气象出版社, 24-31.
|
余世孝, 张宏达, 王伯荪. 1993. 海南岛霸王岭热带山地植被研究(Ⅰ):永久样地的设置与群落类型. 生态科学, 13(2): 13-18. |
臧润国, 刘静艳, 董大方. 1999. 林隙动态与森林生物多样性. 北京: 中国林业出版社.
|
臧润国, 杨彦承, 蒋有绪. 2001. 海南岛霸王岭热带山地雨林群落结构及树种多样性的研究. 植物态学报, 25(3): 270-275. |
臧润国, 余世孝, 刘静艳, 等. 1999. 海南岛霸王岭热带山地雨林林隙更新规律研究. 生态学报, 47(2): 151-158. |
Denslow J S. 1995. Disturbance and diversity in tropical rain forests:the density effect. Ecological application, 5(4): 962-968. DOI:10.2307/2269347 |
Denslow J S. 1987. Tropical rain forest gaps and tree species diversity. Annual review of ecology and systematics, 18: 431-451. DOI:10.1146/annurev.es.18.110187.002243 |
Everham III E M and Brokaw N V, Brokaw N V L. 1996. Forest damage and recovery from catastrophic wind. The botanical review, 62(2): 113-185. DOI:10.1007/BF02857920 |
Lertzman K P, Sutherland G D, Inselberg A and Saunders S C. 1996. Canopy gaps and the landscape mosaic in a coastal temperate rain forest. Ecology, 77(4): 1254-1270. DOI:10.2307/2265594 |
Martínez-Ramos M, Sarukhán J, Pin~ero D. 1988. The demography of tropical trees in the context of forestgap dynamics. London: Blackwell Scientific Publications, 293-313.
|
Moloney K and Levin S A. 1996. The effects of disturbance architecture on landscape-level population dynamics. Ecology, 77(2): 375-394. DOI:10.2307/2265616 |
Pickett S T A, White P S eds. 1985. The ecology of natural disturbanace and patch dynamics. New York: Academic Press.
|
Valverde T and Silvertown J. 1997. Canopy closure rate and forest structure. Ecology, 78(5): 1555-1562. DOI:10.1890/0012-9658(1997)078[1555:CCRAFS]2.0.CO;2 |
Van der Maarel E, Nosest V, Palmer M W. 1995. Variation in species richness on small grassland quadrate:niche structure or small scale plant mobility?. Journal of vegetation science, 6: 741-752. DOI:10.2307/3236445 |
Watt A S. 1947. Pattern and process in the plant community. Journal of ecology, 35: 1-22. DOI:10.2307/2256497 |
Whitmore T C. 1989a. Canopy gaps and the two major groups of forest trees. Ecology, 70(3): 536-538. DOI:10.2307/1940195 |
Whitmore T C. 1989b. Changes over twenty-one years in the Kolombangara rain forests. Journal of ecology, 77: 469-483. DOI:10.2307/2260763 |
Whitmore T C. 1978. Gaps in the forest canopy. New York: Cambridge University Press, 639-655.
|