 2003, Vol. 39
2003, Vol. 39文章信息
- 张莉, 续九如.
- Zhang Li, Xu Jiuru.
- 水分胁迫下刺槐不同无性系生理生化反应的研究
- STUDIES ON PHYSIOLOGICAL AND BIOCHEMICAL RESPONSES OF ROBINIA PSEUDOACACIA CLONES UNDER WATER STRESS
- 林业科学, 2003, 39(4): 162-167.
- Scientia Silvae Sinicae, 2003, 39(4): 162-167.
-
文章历史
- 收稿日期:2000-03-17
-
作者相关文章


2. 北京林业大学森林资源与环境学院 北京 100083
2. College of forest resources and environment, Beijing Forestry University Beijing 100083
当前,水分亏缺条件下植物的生长发育、生理生化、分子生物学及遗传特性的研究愈来愈引起研究者们的关注和重视。迄今,对造林树种的抗旱性研究已开展了大量工作,主要从叶片形态解剖结构、生物量、水分状况、光合特性、蒸腾、细胞膜透性、活性氧代谢、内源激素、渗透调节物质(可溶性糖、脯氨酸等)等方面研究。水分亏缺引起多种生理生化指标的变化,如细胞膜透性、膜脂相变,MDA、Pro含量,SOD、POD、CAT活性等的变化,许多学者已对这些方面进行了有益的探索,但在刺槐无性系上这一研究还未见报道。本研究以刺槐无性系为试材,探讨了水分胁迫下一些生理生化指标的变化规律,以及这些指标间的相互关系,从而说明不同无性系对水分胁迫的反应差异,试图为刺槐的抗旱鉴定及其抗性育种提供理论依据。
1 材料与方法 1.1 试验材料以山东林业厅选育出的10个刺槐(Robinia pseudoacacia)优良无性系即鲁刺1号(Shandongensis 1, S1)、鲁刺2号(Shandongensis 2, S2)、鲁刺7号(Shandongensis 7, S7)、鲁刺13号(Shandongensis 13, S13)、鲁刺42号(Shandongensis 42, S42)、鲁刺59号(Shandongensis 59, S59)、鲁刺68号(Shandongensis 68, S68)、鲁刺78号(Shandongensis 78, S78)、鲁刺102号(Shandongensis 102, S102)、鲁刺箭杆(Shandongensis Jiangan, SJ)为试材。1998年春季进行截杆移根栽植在盆中,土壤为苗圃熟土,按比例配一定细沙。每个品系重复4次,所有苗木按完全随机区组摆放。苗木萌发展叶后按时浇水,并施用有机复合肥料。苗木经过一段时间的缓苗,待长势旺盛于7月份进行控水试验。
1.2 试验方法土壤干旱处理 主要采用自然干燥法得到控水梯度。苗木全部置于温室中以保证外界温湿度环境较一致和防止自然降水的干扰。连续充分浇水5 d后不再浇水,令其自然干燥,分别于断水后24 h,72 h,120 h,168 h取叶样进行各指标的测定。
指标测定 叶组织含水量:烘干法(105℃,8 h);束缚水、自由水含量:折射仪法[阿贝折射仪WAY(2WAJ)];游离脯氨酸:参照朱广廉(1990)编的《植物生理学实验》;可溶性糖含量:蒽酮比色法,620 nm比色;可溶性蛋白含量:考马斯亮蓝染色法,595 nm比色。
2 结果与分析 2.1 土壤干旱对苗木水分状况的影响(1) 苗木叶组织含水量与断水时间的关系 植物组织含水量的高低是决定植物吸水能力大小的一个指标,植物组织含水量越高,表明其吸水能力越弱,在干旱条件下抗旱能力较差。一般说来随着干旱胁迫的加强,植物的组织含水量呈下降趋势。为了解刺槐无性系苗木的水分变化规律,本实验选择不同的断水时间测定其叶组织含水量,测定结果见表 1。
|
|
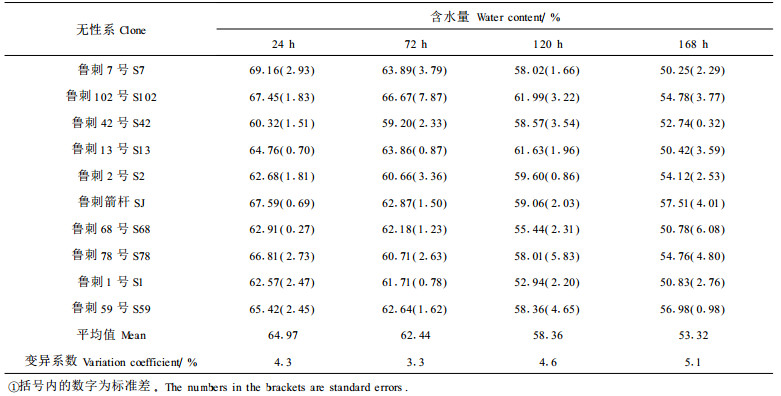
结果表明,供水较正常时,刺槐各无性系苗木的叶组织含水量差别不大,叶片含水量均随断水时间的延长而下降,且不同无性系的下降幅度不同。从表中可知,土壤干旱初期苗木叶片含水量均下降较少,水分状况基本保持稳定,但随着水分胁迫的加剧,所有苗木的叶含水量都迅速下降。干旱初期土壤含水量下降较大,而叶片含水量变化较小是林木对干旱胁迫的适应能力,对刺槐的耐旱具有重要作用。其中无性系鲁刺7号下降最快,下降幅度也最大,总含水量下降27.3%;其次是鲁刺13号,含水量下降了22.1%;而下降最少的是鲁刺42号,仅为12.6%;鲁刺59号仅下降了12.9%,鲁刺2号下降了13.7%。这表明鲁刺42号、鲁刺59号、鲁刺2号对干旱的适应能力较强。同时说明刺槐种内存在水分状况上的差异,这种差异是否为遗传上的差异有待进一步作遗传分析。
(2) 苗木自由水、束缚水含量和束/自与断水时间的关系 马克西莫夫曾指出束缚水数量的增多可决定亲水胶体数量的增多,导致抗旱性增强(蔡玉成等, 1988)。由于束缚水是亲和细胞胶体的水,不易蒸腾,甚至在高温下也难以散失,所以束缚水含量的高低可作为鉴别植物抗旱性的重要指标之一(刘家琼, 1981)。而植物体内束/自常与植物的生长有关,自由水较多时,植株代谢旺盛,生长较快,但抗性弱,束缚水增多使抗逆能力相应提高。同时束/自反映了植物贮水能力的高低,因而国内外学者均采用自由水和束缚水含量及束/自作为主要的耐旱指标。本实验分别测定了不同土壤干燥程度下的苗木自由水和束缚水含量,测定结果见表 2。
|
|
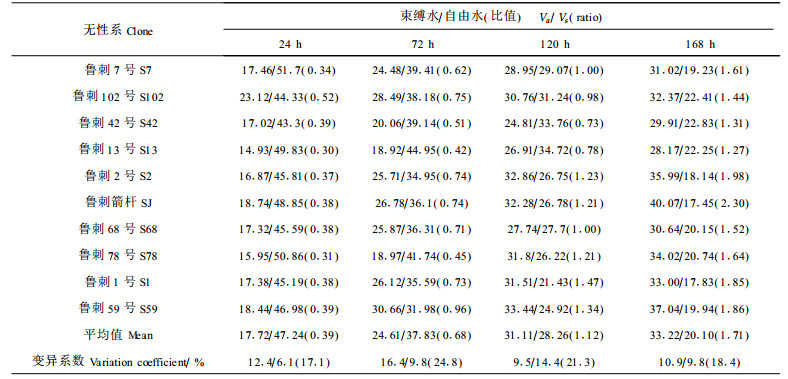
结果表明,当土壤含水量较高时,植株体内的自由水含量均较高,从而表现出较小的束/自比值,此时植物还处于正常生长状态。但随着断水时间的延长,各无性系的自由水含量均呈下降趋势,而束缚水含量则呈上升趋势,从而导致了束/自比值的增高。因此可以看出刺槐无性系在干旱胁迫过程中是以某一种方式来适应的,伴随干旱的加剧其抗旱性逐渐加强。从测定结果得知:在断水168 h,即土壤最干旱时鲁刺箭杆的束/自比值最大,为2.30;其次是鲁刺2号,为1.98;束/自比值最小的是鲁刺13号,为1.27。鲁刺箭杆的束/自比值提高了1.92,鲁刺2号提高了1.61,鲁刺1号、鲁刺59号和鲁刺7号都提高了1.47,而鲁刺102号和鲁刺42号都仅提高0.92。束/自比值的提高幅度反映了各个无性系的抗旱能力大小。从不同无性系束/自比值的变化幅度推测,依靠增加亲水胶体数量来提高自身抗旱能力的途径在无性系间存在差异,而各无性系固有的水分分配状况并不存在差别。这同时也说明植物的抗旱性是在干旱胁迫这一特殊环境中才表现出来的,抗旱基因的表达是受环境因素制约的,进一步证实了树体是基因和环境的统一。
2.2 水分胁迫对苗木游离脯氨酸含量的影响脯氨酸在植物细胞中主要起渗透调节作用,叶片脯氨酸含量增加,可提高细胞溶质含量,增强其渗透调节能力,为此,曾有人提出以干旱时植物体内游离脯氨酸的积累能力作为选育耐旱植物的指标(汤章城, 1983a;1983b)。在不同的土壤干旱条件下,不同刺槐无性系苗木体内游离脯氨酸含量的测定结果见图 1。

|
图 1 苗木叶片游离脯氨酸含量与断水时间的关系 Fig. 1 Relations between free proline content in leaves of seedlings and time of stopping watering |
结果表明,随着土壤干旱程度的加剧,各无性系体内的游离脯氨酸含量均显著增加,而且急剧增加出现时的土壤干旱阈值不同。一般认为植物体内的游离脯氨酸含量不高,只有在水分胁迫时才迅速增加,然而对游离脯氨酸累积的生理效应有多种解释,其与抗旱性的关系也有相反的观点(汤章城, 1983a;1983b)。本实验结果也证明了刺槐无性系在水分胁迫下有迅速的游离脯氨酸积累,其中鲁刺1号、鲁刺59号、鲁刺42号、鲁刺78号积累较多,而鲁刺箭杆积累最少,鲁刺13号次之。在断水72 h时鲁刺13号、鲁刺78号、鲁刺1号就有较明显的累积趋势,当断水120 h时,鲁刺1号、鲁刺13号、鲁刺102号已有大量的积累,而鲁刺7号和鲁刺59号的大量积累则出现在断水168 h时,这说明不同的刺槐无性系对干旱的反应不同,它们适应干旱的主导途径可能有所不同。由于游离脯氨酸累积量的高低除受遗传上的控制外,还受一些外界因素如干旱程度、氮素营养、碳水化合物供给、氨基酸转移的影响,并且不同无性系对这些因素的反应也不同。所以曾有人提出以干旱胁迫下游离脯氨酸大量积累的水势阈值作为评价抗旱能力的重要参考(张殿忠等, 1990),从本研究结果来看具一定的可行性。从测定结果还可看出,水分供应较充足时,刺槐无性系的游离脯氨酸含量都很低,且无性系间差别不大,这可能是游离脯氨酸作为细胞内的一种渗透调节物质对刺槐的渗透调节能力贡献不大的原因,但在干旱逆境中其对稳定渗透调节能力的贡献相对较大。依据游离脯氨酸的累积量可以初步确定无性系中抗旱能力较强的为鲁刺59号、鲁刺1号、鲁刺78号、鲁刺42号。
2.3 水分胁迫对苗木可溶性糖含量的影响在干旱条件下增强细胞渗透调节能力的关键是细胞内渗透调节物质的主动积累。已有研究证明,在细胞的几种渗透调节物质中,对稳定渗透调节能力的相对贡献是K+>可溶性糖>其他游离氨基酸>Ca+>Mg+>脯氨酸(Pro)。所以有人指出在干旱条件下,细胞内渗透调节物质特别是K+和可溶性糖的积累是反映抗旱性强弱的有效指标之一(霍仕平等, 1995)。为此,对不同刺槐无性系苗木在土壤干旱条件下的可溶性糖含量进行了测定,其结果见图 2。

结果表明,所有苗木的可溶性糖含量均随干旱胁迫程度的加强而明显增多,并且增加的幅度在无性系间存在差异。从测定结果显而易见,只有当断水168 h后各无性系苗木的可溶性糖含量才有显著的上升,因此可溶性糖的增加与胁强存在一定的正相关性。这和Al Hakimi等(1995)的叶片中可溶性糖的含量与RWC和胁迫程度呈正相关的研究结论是一致的。当土壤干旱程度较大时,无性系鲁刺13号、鲁刺78号、鲁刺箭杆的可溶性糖含量和积累量都较高,含量分别为10.818,10.392,10.023 g·(100g)-1FW,积累量分别为10.495, 9.623,9.599 g·(100g)-1FW;而鲁刺2号、鲁刺1号、鲁刺59号、鲁刺102号的可溶性糖含量和积累量都较低,含量分别为6.893,6.950,7.238,7.358 g·(100g)-1FW,积累量分别为6.190, 6.672,6.833,6.400 g·(100g)-1FW,这说明不同无性系对干旱的适应性不同。一般说来应以可溶性糖的累积速率来判断植物的抗旱性强弱。从本实验结果得知鲁刺13号积累了97.0%,因此其抗旱性最强;其次是鲁刺箭杆,积累了96.0%,抗旱性次之;第三是鲁刺1号,积累96.0%,抗旱性也较强。积累量较少的鲁刺102号则积累86.7%,鲁刺2号积累89.8%,鲁刺78号积累92.3%,从一侧面说明可溶性糖的主动积累在这些无性系上反应较不明显,其渗透调节能力可能较弱。可溶性糖的大量累积存在一个水势阈值,从实验结果来看,不同刺槐无性系的可溶性糖积累的水势阈值差别不大,并且其积累与胁迫时间有一定的相关性。可溶性糖的缓慢积累是刺槐无性系对水分胁迫的一种适应性反应,糖量增加均达到原来含量的7倍以上,说明可溶性糖可能是刺槐无性系渗透调节的主要物质。
2.4 水分胁迫对苗木可溶性蛋白含量的影响很少有研究注重随干旱强度加大,植物组织通过可溶性物质的主动积累来降低渗透势,进行渗透调节。而不同植物溶质积累程度的不同,根本原因是它们的基因表达有区别,基因表达(转录和翻译)的产物是蛋白质,研究干旱胁迫下蛋白质的变化规律是研究抗旱基因的重要基础,它与抗旱性存在某种关系。已有大量研究证实干旱抑制蛋白质的合成并诱导蛋白质的降解,从而使植株体内的总蛋白质含量降低(魏良民, 1991)。蛋白质含量的降低与植物的衰老密切相关,这也是逆境对植物的一种伤害作用。但有研究表明(于同泉等, 1995),干旱胁迫下可溶性蛋白的含量呈增加趋势,所以蛋白质的代谢变化在植物逆境生理中的意义尚不明确。本实验对刺槐不同无性系在干旱胁迫下可溶性蛋白的含量进行了测定,其结果见表 3。
|
|
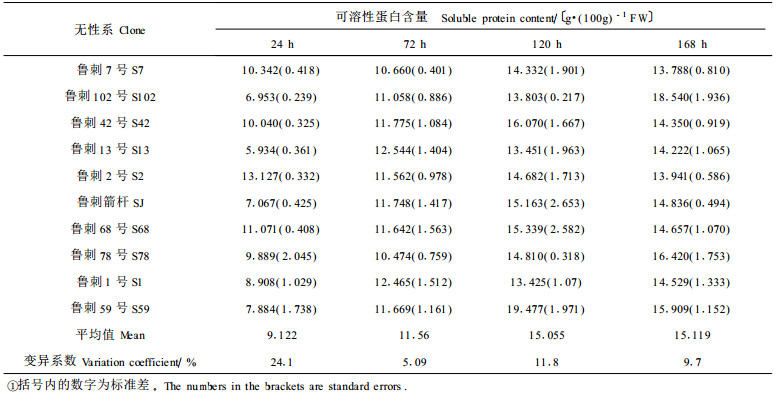
研究结果表明,随着断水时间的延长,刺槐各无性系的可溶性蛋白含量均呈上升趋势,但上升过程中有波动。总的说来,苗木体内可溶性蛋白含量的增加幅度不大,这可能由于干旱对蛋白合成抑制和降解的作用较大。其中,鲁刺102号、鲁刺13号、鲁刺1号的可溶性蛋白含量持续上升,而其余的则在断水120 h时达到顶峰,其后又呈下降趋势,但总水平都有所提高。其中鲁刺102号增加最多,增加了62.5%,鲁刺13号也增加了58.3%,鲁刺箭杆增加52.4%,而鲁刺2号仅增加5.8%,鲁刺68号增加24.5%,鲁刺7号增加25.0%。这种不同的变化幅度可能是不同无性系对干旱的适应性反应差异造成的。有研究表明,水分胁迫下可溶性蛋白的增加说明这些蛋白在增强抗旱性上起重要作用,研究者认为可溶性蛋白水平的提高有渗透调节和延缓被干旱胁迫加速的衰老过程的作用1)。本研究还证明随蛋白含量的增加,其他渗透调节物质如可溶性糖、游离脯氨酸等亦随之增加。这是因为植物的抗旱能力是由基因表达所控制的,表达的最终效应是一系列的生理生化方面的变化。所以可以猜测有新生蛋白存在,它们是水分胁迫的产物,称为“旱胁蛋白”。至于该研究的新增蛋白的组成还有待于进一步研究。但值得提出,迄今为止,当胁迫强度达到阈值时可溶性蛋白的增加,其来源途径和生理意义尚未明了。
1) 陈绍良.杨树种间耐旱性差异的生理生化基础研究.北京林业大学,博士学位论文,1997
3 结论刺槐不同无性系在土壤干旱时,其组织含水率呈现下降的趋势,但在无性系间下降的幅度有差别。同时在干旱胁迫时,束缚水含量出现上升的趋势,而自由水含量则出现下降趋势,束/自比值明显增大。各无性系的水分分配状况的区别,说明它们适应干旱的途径不同。
刺槐不同无性系在土壤干旱时,各苗木体内的游离脯氨酸含量出现大量的积累,积累量在无性系间存在差异。不同无性系表现出迅速积累的土壤干旱阈值有差异,这是不同无性系对土壤干旱的适应差异的表现。
刺槐不同无性系在土壤严重干旱时,所有苗木都表现出可溶性糖的大量积累,积累量在无性系间存在差异。可溶性糖的迅速积累与干旱胁迫强度与持续时间有一定的正相关性,其可能是刺槐体内的一种重要的渗透调节物质。
刺槐不同无性系在土壤干旱时,体内可溶性蛋白含量也呈现出上升的趋势,但上升的过程中有波动。可溶性蛋白含量增加的幅度都较小,这是由于干旱胁迫使脂质过氧化产生了与蛋白质复合体反应并使之失活的有毒物质而抑制蛋白质的合成或膜蛋白在过氧化过程中受到伤害,从而减少蛋白质含量(陈少裕, 1991)。从可溶性蛋白的增加可以推测有“旱胁蛋白”产生,这种热稳定性蛋白的生成有利于增强苗木的抗旱性。
刺槐不同无性系在土壤干旱时表现出多种生理生化指标的不同变化,单独依靠某一指标来评判不同无性系的抗旱能力大小所得结果有出入,因而需将多项指标综合起来考虑,从而较为全面准确地评价抗旱性强弱。
不同刺槐无性系在干旱胁迫时生理生化指标的不同变化规律及它们间的相关性说明了组织在水分亏缺条件下叶片代谢的变化及其相互关系。这些生理生化变化又是组织对水分胁迫的一种防御、躲避和忍耐的生理机理。以这些指标的变化作为抗旱性鉴定指标,可通过代谢变化来反映刺槐不同无性系的抗旱机理。
蔡玉成, 王政琦, 贾福琴. 1988. 白榆不同地理种源抗旱性的比较. 甘肃林业科技, (2): 6-11. |
陈少裕. 1991. 膜脂过氧化对植物细胞的伤害. 植物生理学通讯, 27(2): 84-90. |
霍仕平, 晏庆九, 宋光英, 等. 1995. 玉米抗旱鉴定的形态和生理生化指标研究进展. 干旱地区农业研究, 13(3): 67-73. DOI:10.3321/j.issn:1000-7601.1995.03.003 |
刘家琼. 1981. 柠条、花棒生理特性的研究. 林业科技通讯, (6): 6-8. |
汤章城. 1983a. 植物对水分胁迫的反应和适应性. 植物生理学通讯, 19(3): 24-29. |
汤章城. 1983b. 植物对水分胁迫的反应和适应性. 植物生理学通讯, 19(4): 1-7. |
魏良民. 1991. 几种旱生植物碳水化合物和蛋白质变化的研究. 干旱区研究, 8(4): 38-41. |
于同泉, 柴丽娜, 刘宗萍. 1995. 水分胁迫下小麦幼苗可溶性蛋白质的表现与小麦抗旱蛋白之初探. 北京农学院学报, 10(1): 26-30. |
张殿忠, 汪沛洪, 席连喜. 1990. 干物质累积和脯氨酸累积的水势阈值与小麦抗旱性的关系. 干旱地区农业研究, 8(2): 66-71. |
朱广廉, 钟海文, 张爱琴. 1990. 植物生理学实验. 北京: 北京大学出版社.
|
Al Hakimi A, Monneveux P, Galiba G. 1995. Soluble sugars, proline, and relative water content (RWC) as traits for improving drought tolerance and divergent selection for RWC from T. polonicum to T. durum. J of Genetics & Breeding, 49(3): 237-243. |