 2003, Vol. 39
2003, Vol. 39文章信息
- 李国旗, 安树青, 张纪林, 李瑾, 王云静.
- Li Guoqi, An Shuqing, Zhang Jilin, Li Jin, Wang Yunjing.
- 土壤盐胁迫下杨树次生木质部的解剖特征
- THE ANATOMICAL CHARACTERISTICS OF POPULUS DELTOIDES(LUX) UNDER STRESS OF VARIOUS SOIL SALINITIES
- 林业科学, 2003, 39(4): 89-97.
- Scientia Silvae Sinicae, 2003, 39(4): 89-97.
-
文章历史
- 收稿日期:2002-04-08
-
作者相关文章



2. 江苏省林业科学研究所 南京 211153;
3. 南京大学生物科学与技术系 南京 210093
2. Jiangsu Forestry Science Research Institute Nanjing 211153;
3. Biological Science & Technology Department of Nanjing University Nanjing 210093
杨树(Poplar)属于杨柳科(Salicaceae),广泛生长于北半球。世界上天然杨树林的面积为2×107 hm2左右,主要分布于美国和加拿大之间。国外杨树人工林面积总和只有1.4×106 hm2, 主要分布在法国、西班牙、意大利和韩国等国家;而中国的杨树不仅品种多,而且绝大多数为人工林,面积达到6.6×106 hm2(王世绩,1995)。大面积的杨树人工林极大地缓解了我国木材需求的压力。杨树生长快、耐盐碱和瘠薄,不仅可以栽植在不宜农用的地块中,而且可以和农作物进行多种形式的间作。杨树以其极大的经济意义常常被称为多年生的木本作物(FAO, 1979)。
中国是一个拥有1.8×104 km大陆海岸线的海洋大国,同时也是世界上海洋灾害最为严重的国家(杨华庭, 1992; 高建华等,1999)。在我国国民经济发展中占有举足轻重地位的沿海地区,每年遭受的海洋灾害损失非常巨大,尤其是台风所引起的直接经济损失增长最快,年平均损失从1950年的不足1亿元,上升到1990年的100亿元以上,1997年高达300亿元(汪纬林等,1999)。自从实施沿海防护林工程以后,我国沿海地区营造了大量的杨树人工林,成为沿海地区工农业生产的安全屏障,在防风减灾、保持水土、调节气候等方面发挥积极作用,是维持沿海地区的生态系统平衡、实现可持续发展的必要条件(季子修等,1996)。但是处于海洋灾害最前沿的沿海防护林,所遭受的盐胁迫最直接、最严重。基于全球气候变暖、海平面上升、海岸防护林遭受的盐胁迫将更加严重(吴崇泽,1994;赵希涛等,1992;杨桂山等, 1999)。本文对海岸带地区不同土壤含盐量下的杨树进行了解剖,旨在从微观角度揭示盐胁迫对杨树材质的影响机制,从而有助于筛选耐盐的杨树品系,为抗风杨树的诊断和选育提供参考指标。
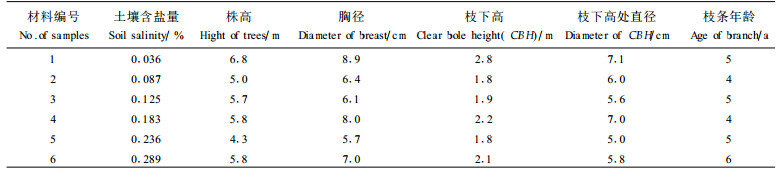
1 材料和方法 1.1 实验材料在江苏省如东县东凌乡的海岸防护林带4~6 a生人工杨树林中, 经过初选和复选,选择了6个不同梯度土壤含盐量的样点,每个样点选择2株杨树,均在枝下高处朝南的方向截取一段健康的枝条,在取枝条样品的同时,再取土壤样品,测得6个样点的土壤含盐量,见表 1。在垂直海堤2 km的取样地段,地形、地貌及各种气象因子都一样,在影响杨树生长的各种生态因子中,土壤含盐量是最重要的影响因子。有资料表明,杨树能忍耐的含盐量是0.3%,极限含盐量是0.34%(杨雄鹰等,1995)。由此可见,东凌乡的土壤虽然按盐碱程度分类为轻中度盐碱地,而对杨树来说,盐胁迫还是相当严重的。
|
|
按张新英等(1990)的方法制作切片,经过预实验,离析材料的染色用代氏苏木精染料代替星蓝,用中性树胶代替冷杉胶和upper胶。染色液的配方按李正理(1987)的方法配制。
材料切片 将木材的边材部分切成1 cm3的小块,水煮软化后在Histoslide-2000滑走切片机上切横、径、弦3个切面的切片,厚度约16 μm。用番红和代氏苏木精的混合液染色,酒精脱水,用中性树胶封片。
染色和封片 样品放入50%酒精中→番红苏木精(体积比为9:1)→1 h后转入70%酒精→用80%的酒精漂洗→用90%的酒精漂洗→95%的酒精漂洗→用纯酒精漂洗→用二甲苯透明→用中性树胶封片。
材料离析 将木材切成1 cm长,3 mm×3 mm的长条,用过氧化氢和冰醋酸的混合液离析,代氏苏木精染色,中性树胶封固。
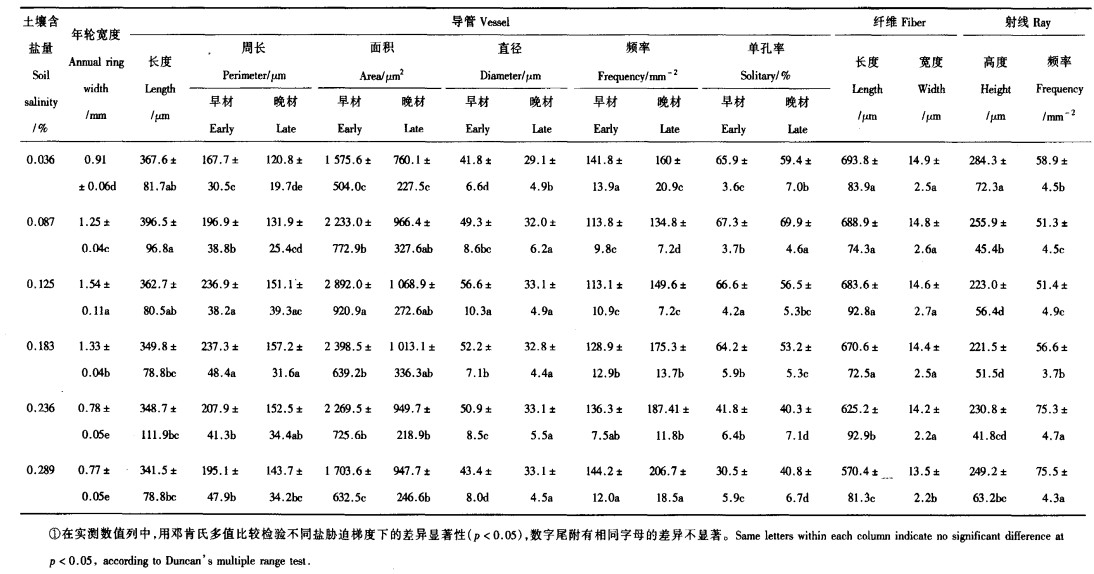
1.2.2 解剖指标的测定用XY-540型彩色图像分析仪测定早材和晚材导管分子直径、早材和晚材导管频率、纤维长度和纤维宽度。其中,横切面上的导管频率测定10个视野;其它每个指标均测量50次,然后计算平均值和方差。纤维的长度和宽度在材料离析中测量,其它指标均在切片中测量。
根据1989年国际木材解剖学家协会(IAWA committee)制定的“阔叶树材识别显微特征表”(杨家驹等,2002)和“中国木材志”(成俊卿等,1992)所用术语,对杨树的形态特征进行描述。
2 结果与分析 2.1 盐胁迫对杨树次生木质部形态特征的影响杨树的次生木质部有导管、纤维(轴向系统)和射线(径向系统)组成(李正理等,1983)。在不同的土壤盐度胁迫下,杨树的次生木质部的组成分子趋于小型化,形态特征的差别不明显,而数量特征的差别比较显著(表 2)。
|
|
杨树属于散孔材,生长轮不很明显。导管横切面为卵圆及椭圆形,略具多角形轮廓。径列复管孔(多为2~4)及单管孔(图版Ⅰ-1,2)。管间纹孔式互列,多角形;单穿孔,圆形、圆卵形及椭圆形,穿孔板略倾斜至倾斜(图版Ⅰ-3,4)。导管壁厚1~2 μm, 其端壁倾斜并具有长尾。导管有具缘纹孔,射线间的纹孔为半具缘纹孔,未见侵填体。轴向薄壁组织量少,呈轮界状、稀星散状,未见晶体。木纤维壁薄,木射线非叠生,射线单列,高3~15个细胞(图版Ⅰ-4);射线类型为同型单列射线(图版Ⅰ-5,6)。

|
图版Ⅰ Plate Ⅰ 1和2分别为0.036%和0.289%土壤盐度下的横切面, 示散孔材, 7x;3和4分别为0.036%和0.289%土壤盐度下的弦切面, 33×;5和6分别为0.036%和0.289%土壤盐度下的径切面, 66x。 1 and 2:Transverse sections on various soil salinity, 0.036%and 0.289%, respectively, showing wood diffuse-porous, 7×; 3 and 4:Radial section on vari-ous soil salinity, 0.036%and 0.289%, respectively, 33×; 5 and 6:Tangential section on various soil salinity, 0.036%and 0.289%, respectively, 66x. |
当土壤含盐量从0.036%→0.087%→0.125%升高时,杨树的年轮平均宽度(ARW)随含盐量的升高而升高,0.91→1.25→1.54 mm,但是土壤含盐量继续从0.125%→0.183%→0.236%→0.289%升高时,ARW随含盐量的升高而降低,1.54→1.33→0.78→0.77 mm, 可见0.125%的含盐量是ARW变化的临界点, 证明盐胁迫是存在的。从0.036%上升到0.289%,土壤含盐量增加7.03倍,杨树的年轮宽度减小了15.4%,方差检验表明ARW差异显著,可见土壤含盐量明显影响了杨树的生长(图 1)。

|
图 1 不同土壤含盐量下杨树年轮和导管长度的变化 Fig. 1 Variation of annual ring width and vessel length of Poplar in various soil salinities |
(1) 对导管长度的影响 在盐胁迫条件下,导管长度的变化曲线与年轮的变化曲线极为相似(图 1)。当土壤含盐量从0.036%→0.087%升高,杨树的导管长度(VL)也从367.6→396.5 μm,而当土壤含盐量继续从0.087%→0.125%→0.183%→0.236%→0.289%升高时,VL却从396.5→362.7→349.8→348.7→341.5 μm逐渐降低,可见0.087%的含盐量VL变化的临界点。但是,方差检验表明VL的增加值和下降值之间都没有显著差异(表 2)。(2)对面积、周长和直径的影响 由于早材的导管大于晚材的导管,因此早材导管的面积、周长和直径均大于晚材的面积、周长和直径(图 2、3、4)。早材和晚材的导管面积(VA)、周长(VP)和直径(VD)均表现为先升高后降低。当土壤含盐量从0.036%→0.087%→0.125%升高时,导管面积(VA)在早材和晚材中都升高,1 575.6→2 233.0→2 892.0 μm2和760.1→966.4→1 068.9 μm2;而土壤含盐量继续从0.125%→0.183%→0.236%→0.289%升高时,早材和晚材的VA都开始减小,2 892.0→2 398.5→2 269.5→1 703.6 μm2和1 068.9→1 013.1→949.7→947.7 μm2,可见0.125%的土壤含盐量是VA变化的临界点。土壤含盐量从0.036%→0.087%→0.125%→0.183%升高时,导管周长(VP)在早材和晚材都升高,167.7→196.9→236.9→237.3 μm和120.8→131.9→151.1→157.2 μm;含盐量继续从0.183%→0.236%→0.289%升高,早材和晚材的VP减小,分别为237.3→207.9→195.1 μm和157.2→152.5→143.7 μm,可见0.183%的土壤含盐量是VP变化的临界点。含盐量从0.036%→0.087%→0.125%,导管直径(VD)在早材和晚材中的增加分别为41.8→49.3→56.6 μm和29.1→32.0→33.1 μm,含盐量继续升高0.125%→0.183%→0.236%→0.289%,VD的变化分别为56.6→52.2→50.9→43.4 μm和33.1→32.8→33.1→33.1 μm,可见0.125%的土壤含盐量是导管直径的临界点。上面的结果说明,面积和直径对土壤含盐量的反应较周长敏感,当含盐量大于0.125%时,盐胁迫使导管变短、变粗,有利于水分的吸收。导管分子的直径、周长和面积是3个密切相关的指标,它们在度量导管的输导水分有效性方面,具有同等作用,而3个指标的变化趋势完全相同,正好起到相互验证的作用。早材和晚材的VA、VP和VD 6个指标都随着土壤含盐量的升高而增加,达到临界点以后,都开始减小。方差分析表明,除了晚材的VA和VD两个指标以外,其余4个指标在临界点的值均与其降低值之间有显著差别(表 2)。(3)对导管频率的影响 当土壤含盐量从0.036%→0.087%→0.125%→0.183%→0.236%→0.289%升高时, 导管频率(VF)在早材和晚材中的变化分别为141.8→113.8→113.1→128.9→136.3→144.2 mm-2和160→134.8→149.6→175.3→187.41→206.7 mm-2,二者均表现为先降低后升高,这与导管直径的变化正好相反(图 5)。早材和晚材VF变化的临界点分别为0.125%和0.087%。方差检验表明,随着含盐量的升高,早材和晚材中导管频率之间都有显著差异(表 2)。(4)对导管单孔率的影响 在不同梯度的含盐量条件下,早材的单孔率均比晚材高;早材和晚材的单孔率(PSV)均表现为先升高后降低的趋势,PSV的降低表明受盐胁迫后导管趋向于聚集分布(图 6)。在6种含盐量梯度0.036%→0.087%→0.125%→0.183%→0.236%→0.289%下,早材和晚材中PSV的变化分别为65.9%→67.3%→66.6%→64.2%→41.8%→30.5%和59.4%→69.9%→56.5%→53.2%→40.3%→40.8%。方差分析表明,当土壤含盐量升高时,早材和晚材的导管单孔率的变化均达到显著水平(表 2)。

|
图 2 不同土壤含盐量下杨树导管面积的变化 Fig. 2 Variation of vessel area of Poplar in various soil salinities |

|
图 3 不同土壤含盐量下杨树导管周长的变化 Fig. 3 Variation of vessel perimeter of Poplar in various soil salinities |

|
图 4 不同土壤含盐量下杨树导管直径的变化 Fig. 4 Variation of vessel diameter of Poplar in various soil salinities |

|
图 5 不同土壤含盐量下杨树导管频率的变化 Fig. 5 Variation of vessel frequence of Poplar in various soil salinities |

|
图 6 不同土壤含盐量下杨树导管单孔率的变化 Fig. 6 Variation of solitary vessel of Poplar in various soil salinities |
当土壤含盐量从0.036%→0.087%→0.125%→0.183%→0.236%→0.289%升高, 杨树木质部中的纤维长度(FL)和纤维宽度(FW)的变化分别为693.8→688.9→683.6→670.6→625.2→570.4 μm和14.9→14.8→14.6→14.4→14.2→13.5 μm,FL和FW均减小(图 7)。方差分析表明,只有在土壤盐度达到或大于0.183%时,FL和FW的减小才达到显著水平(表 2)。

|
图 7 不同土壤含盐量下杨树纤维长度和宽度的变化 Fig. 7 Variation of fiber length and fiber width of Poplar in various soil salinities |
在6个土壤盐度0.036%→0.087%→0.125%→0.183%→0.236%→0.289%下, 木射线高度(RH)和射线频率(RF)的变化分别为284.3→255.9→223.0→221.5→230.8→249.2和58.9→51.3→51.4→56.6→75.3→75.5(图 8)。0.125%的土壤含盐量是RH和RF由降低到升高的临界点。射线高度减小是由于生长受到抑制、年轮宽度减小所致,射线频率的增加意味着横向运输效率的提高。方差分析表明,在早材和晚材中,RH和RF的下降和升高都是显著的(表 2)。

|
图 8 不同土壤含盐量下杨树射线高度和频率的变化 Fig. 8 Variation of ray height and ray frequency of Poplar in various soil salinities |
在微观水平上,能够表征杨树生长的主要解剖指标有年轮宽度、纤维长度和宽度、射线高度。邓亮等(1989)的研究表明:随着海拔的升高(降雨量增加),紫萼丁香(Syringa oblata var.giraldii)的年轮宽度、纤维长度和宽度、射线高度都增加,并且年轮宽度与海拔高度之间存在显著的线性相关关系。Villar等(1997)也得到了类似的结果,证明在不同气候梯度下栎属(Quercus)3个树种的木质部特征与温度无关而与降雨量有关系。可见在众多的环境影响因子中,降雨量是树木上生长最主要的限制因子之一。实际上,水分亏缺是大多数非生物胁迫的共同属性,盐胁迫和低温胁迫也都是由于水分亏缺而引起植物受害(王伟,1998)。本项研究发现,尽管在6种土壤盐度下,年轮宽度、纤维长度、纤维宽度和射线高度的变化表现不一致,但是有2点是共同的:首先,临界点之后,随着土壤盐度的升高,年轮宽度、纤维长度、纤维宽度和射线高度均表现为减小的趋势,这说明随着土壤盐度的增加,杨树的生长受到抑制;其次,年轮宽度、纤维长度、纤维宽度和射线高度的变化曲线都存在拐点,这些拐点的变化范围在土壤盐度为0.125%~0.183%之间,这些说明了杨树的不同解剖指标对土壤盐度的敏感性是不同的。由于盐度能够改变细胞壁的伸展性能,使细胞壁的生长受到抑制,从而使生长受到抑制(Neumann,1993)。至于从更微观的层次来探讨盐胁迫如何使杨树的材质发生变化,已有的研究表明:渗透调节是杨树耐盐性的重要生理基础之一,因为耐盐性强的胡杨(Populus euphratica)叶和根中含有较高的甜菜碱和糖类物质,而耐盐性弱的群众杨(P.popularis)则较低(陈少良等,2001)。盐胁迫后植物体内会产生大量的过氧化物,从而造成伤害,过氧化物酶(POD)能够消除过氧化物和活性氧的伤害(廖祥儒等,1996)。此外,在许多木本植物中,POD是一种重要的IAA侧链氧化酶(Harkin et al., 1973),它不仅通过调节生长素(IAA)水平参与形成层活动的控制,而且直接参与作为细胞壁的3大组成成分之一的木质素的合成(Reineicke et al., 1987)。
3.2 盐胁迫对杨树水分输导的影响水分的运输是从高势能区向低势能区进行的,一般情况下,植物上部水势比下部低,故水分流动从下到上。与水分流动有关的参数很多,其中以导管直径和导管频率最为重要。从某种角度上讲,导管直径与水分输导的有效性有关,导管频率与水分输导的安全性直接相关(Bass, 1982)。木质部中水分的流率与导管直径的4次方成正比(林金星等,1993),即导管直径越大,导管的输导率越高。假如一种植物有3种大小不同的导管直径:40、80、160 μm,它们的相对直径分别是1,2和4,在可比条件下,流率分别为1,16和256。显然,在同一植株内,160 μm的大导管运输了93.8%的水分;而直径为40 μm的导管只运输了水分的0.4%。Zimmermann(1983)把大导管高流率的现象,称为“有效性”。有效性高并不意味着植物都会向大导管方向进化,因为大导管在提高输导有效性的同时,影响了输导的安全性。众所周知,导管并不是理想的毛细管,导管具有不同的纹孔结构,而且导管分子长短不一,其输导效率取决于多因子的综合作用。一旦遭到虫害袭击或外界的机械破坏,空气进入导管,引起输导系统阻塞。大导管的植株在某一断面出现阻塞的可能性比小植株高256倍。冬季,大导管内更容易出现冰冻,解冻时常出现气泡,这说明大导管可能降低水分输导的安全性。由于导管直径代表水分输导的有效性,导管频率代表水分输导的安全性,当土壤含盐量从0.036%升高到0.289%, 导管分子直径在早材和晚材中分别从41.8 μm增加到43.4 μm,29.1 μm到33.1 μm,增加的百分率分别为3.7%和13.7%,输水效率分别提高16.2%和167.2%。
导管频率的变化为先降低后升高,正好与导管直径变化趋势相反,说明水分输导的有效性和安全性是一对此消彼长的矛盾。如果将导管直径和导管频率的曲线作在一个图中,必然会有两个交叉点,在0.075%(A)和0.23%(B)的土壤含盐量附近,由于A点的土壤盐度很低,能够刺激杨树的生长,导管直径有增大的趋势,也就是输水的有效性提高,但导管频率的降低表明,有效性不能无限扩大,这必然受制于安全性;B点属于中度含盐量,表明杨树受盐胁迫后生长受到抑制,导管频率增加,输水安全性提高,但是安全性的提高也必然受有效性的制约。可见,B点是杨树受盐胁迫后水分输导的有效性和安全性的平衡点。本文结果发现盐胁迫使杨树的导管直径和导管频率增加,这说明适度的盐胁迫使导管输导水分的安全性和有效性都有所提高,因此可以把导管直径和导管频率作为筛选抗风耐盐杨树品种的参考指标之一。植物在水分输导的有效性与安全性之间寻求平衡点,这是植物适应环境的智慧。在长期的进化过程中,植物通过自身的内部调节,来适应环境条件的变化,达到运输有效性和安全性的协调与统一。在潮湿地区,导管较大,频率较低(Bass et al., 1987), 在干旱地区和海滨盐渍环境,树木导管直径变小,频率明显上升,尽管导管直径变小,在一定程度上使水分运输的有效性受到影响,但小导管机械强度大,导管坍塌的可能性也随之减少。
方精云等(2000)的研究结果表明,水分因子(年降水和水分综合指数)是影响亮叶水青冈(Fagjus lucida)形态解剖特征的主导生态因子。随着水分因子的增加,亮叶水青冈的导管直径和导管长度以及纤维长度均呈明显增加趋势,导管频率、射线高度则呈下降趋势。林鹏等(2000)的研究证明,紫金牛科的桐花树(Aegiceras corniculatum)和海桑(Sonneratia caseolaris)在海滨盐渍环境下,导管直径变小、数量增多;而导管频率的增加也使导管横截面积也相应增加,水分运输的有效性也得到一定的补偿。这些结论都与本文的实验结果是一致的。
Carlquist(1984)曾经注意到逆境中导管的聚集现象,提出了复管孔(vessel groupings)的概念。已有的研究结果发现,复管孔是对季节性干旱的适应,沙生植物的导管直径小、复孔率高(Lindorf,1997;张新英等,1993;Roth,1996),这与本文的研究结果是完全一致的。本文结果证明,当土壤含盐量从0.036%升高到0.289%时,杨树的早材和晚材的导管单孔率分别降低了35.4%和18.6%,说明在盐胁迫条件下,杨树导管的复孔率升高,导管呈聚集分布,这样不仅可以增加水分输导,也具有机械支持功能。
4 结论本文对生长在不同土壤盐度下杨树次生木质部的形态特征和数量特征进行了深入研究,解剖表明,在不同的土壤盐度胁迫下:(1)杨树的次生木质部形态特征的变化不明显,但是组成分子趋于小型化。(2)次生木质部数量特征的变化比较明显,即随着土壤含盐量的升高,一些表征生长量的指标(年轮宽度、纤维长度、纤维宽度、射线高度和频率)均显著减小,说明盐胁迫下杨树的生长受到抑制。杨树只有依靠增加水分吸收才能缓解盐胁迫所带来的压力,表现在:代表水分输导有效性指标(导管直径、周长和面积)和安全性指标(导管频率)均增加了,说明杨树是以其导管缩短变粗(导管长度减小而直径增大)、聚集分布(导管单孔率减小)来适应土壤盐胁迫的。
成俊卿, 杨家驹, 刘鹏. 1992. 中国木材志. 北京: 中国林业出版社, 572-586.
|
陈少良, 李金克, 毕望富, 等. 2001. 盐胁迫条件下杨树盐分与甜菜碱及糖类物质变化. 植物学通报, 18(5): 587-596. DOI:10.3969/j.issn.1674-3466.2001.05.011 |
邓亮, 张新英. 1989. 生长在太白山上的紫萼丁香木材的生态解剖. 植物学报, 31(2): 95-102. |
方精云, 费松林, 樊拥军, 等. 2000. 贵州梵净山亮叶水青冈解剖特征的生态格局及主导因子分析. 植物学报, 42(6): 636-642. DOI:10.3321/j.issn:1672-9072.2000.06.016 |
高建华, 朱晓东, 余有胜, 等. 1999. 我国沿海地区台风灾害影响研究. 灾害学, 14(2): 73-77. |
季子修, 施雅风. 1996. 海面上升、海岸带灾害与海岸防护问题. 自然灾害学报, 5(2): 56-64. |
李正理. 1987. 植物制片技术. (第二版). 北京: 科学出版社.
|
李正理, 张新英. 1983. 植物解剖学. 北京: 高等教育出版社, 126-133.
|
廖祥儒, 朱新产. 1996. 活性氧代谢和植物抗盐性. 生命的化学, 16(6): 19-22. |
林金星, 林金安.导管及其在植物水分运输中的作用.见: 林金安主编.植物科学综论.哈尔滨: 东北林业大学出版社, 1993: 125-137
|
林鹏, 林益明, 林建辉. 2000. 桐花树和海桑次生木质部的生态解剖. 林业科学, 36(2): 125-128. DOI:10.3321/j.issn:1001-7488.2000.02.021 |
王世绩主编.杨树研究进展.北京: 中国林业出版社, 1995: 387-391
|
王伟. 1998. 植物对水分亏缺的某些生化反应. 植物生理学通讯, 34(5): 388-393. |
汪纬林, 毛桐恩, 解敬. 1999. 我国天灾综合预测研究进展. 科技导报, (1): 46-48. DOI:10.3321/j.issn:1000-7857.1999.01.014 |
吴崇泽. 1994. 海平面上升对海岸带环境的影响与危害及其防治对策. 灾害学, 9(1): 34-37. |
杨桂山, 施雅风. 1999. 中国海岸地带面临的重大环境变化与灾害及其防御对策. 自然灾害学报, 8(2): 13-19. |
杨华庭.中国海洋灾害基本情况及其防御对策.见: 施雅风, 黄鼎成, 陈泮勤主编.中国自然灾害灾情分析与减灾对策.武汉: 湖北科学技术出版社, 1992: 259-267
|
杨家驹, 衣铁梅译.阔叶树材识别显微特征表.见: 侯元兆编著.热带林学.北京: 中国林业出版社, 2002: 208-273
|
杨雄鹰, 徐维坤, 黄胜利, 等. 1995. I-69杨耐盐力研究. 林业科学研究, 8(3): 348-350. |
张新英, 曹宛虹. 1990. 不同生境下的沙棘次生木质部解剖学的研究. 植物学报, 32(12): 909-915. |
张新英, 曹宛虹. 1993. 豆科7种沙生植物次生木质部的生态解剖. 植物学报, 35(12): 929-935. |
赵希涛, 杨达源主编. 1992. 气球海面变化. 北京: 科学出版社, 156-160.
|
Bass P. 1982. New perspectives in wood anatomy. Nijhoff, Junk. The Hague: 252-263. |
Bass P, Schweigruber F H. 1987. Ecological trends in the wood anatomy of trees, shrubs and climbers from Europe. IAWA Bulletin, 8: 245-276. DOI:10.1163/22941932-90001053 |
Carlquist S. 1975. Ecological stratigies of xylem evolution. Berkley: University of California Press.
|
Food and Agriculture Organization of The United Nations(FAO). 1979. Poplars and Willows. Rome: Published under the auspices of the international poplars commission, 293-302.
|
Harkin J M, John R O. 1973. Lignification in trees: indication of exclusive peroxidase participation. Science, 180: 296-298. DOI:10.1126/science.180.4083.296 |
Lindorf H. 1997. Wood and leaf anatomy in sessea corymbiflora from an ecological perspective. IAWA Journal, 18(2): 157-168. DOI:10.1163/22941932-90001477 |
Neumann P M. 1993. Rapid and reversible modifications of extension capacity of cell wall in elongating maize leaf tissue responding to root addition and removal of NaCl. Plant Cell Environment, 16: 1107. DOI:10.1111/j.1365-3040.1996.tb02068.x |
Reineicke D M, Baandurski R S. Auxin biosynthesis and metabolism. In: Davis P J eds. Plant hormone and their role in plant growth and development. Kiuwen: Academic publishers, 1987: 24-43
|
Roth A. 1996. Water transport in xylem conduits with ring thickenings. Plant cell and environment, 19(5): 622-629. DOI:10.1111/j.1365-3040.1996.tb00397.x |
Villar S P, Castro D P, Perez R C, et al. 1997. Stem xylem features in three Quercus (Fagaceae) species along a climatic gradient in NE Spain. Tree structure and function, 12(2): 90-96. |
Zimmermann M H. 1983. Xylem structure and the ascent of sap. Berlin: Springer.
|