 2003, Vol. 39
2003, Vol. 39文章信息
- 陈金慧, 施季森, 诸葛强, 黄敏仁.
- Chen Jinhui, Shi Jisen, Zhuge Qiang, Huang Minren.
- 杂交鹅掌楸体细胞胚胎发生研究
- STUDIES ON THE SOMATIC EMBRYOGENESIS OF LIRIODENDRON HYBRIDS(L.CHINENSE×L. TULIPIFERA)
- 林业科学, 2003, 39(4): 49-53.
- Scientia Silvae Sinicae, 2003, 39(4): 49-53.
-
文章历史
- 收稿日期:2002-12-12
-
作者相关文章



木兰科(Magnoliaceae)鹅掌楸属(Liriodendron)植物,现仅存2个种,即鹅掌楸(L. chinense)和北美鹅掌楸(L. tulipifera),分隔在中国和北美洲大陆。由于地理隔离,天然状态下,未见有种间杂交种的报道。2个种间杂交种于1963年由我国已故著名林木遗传育种学家叶培忠教授创造,并表现出明显的杂种优势(南京林产工业学院林学系育种组, 1963)。杂交鹅掌楸树形美观,叶形奇特,花大鲜艳,象郁金香,因此是一种重要的庭院绿化树种。另外,据李周歧(2001)、季孔庶(2001)报道,1a生杂交鹅掌楸生长速度比中国鹅掌楸快30%左右,2 a生时可快60%,5 a生时高达100%~200%,杂种优势十分明显。成年鹅掌楸树体高大,可达60 m,也是一种重要的造林树种和用材树种。在美国,北美鹅掌楸木材大量用作装饰材、工业胶合板面材或芯材、造纸等(Merkle et al., 1986)。杂交鹅掌楸虽然有许多优越性,但杂交制种受到季节限制,影响了杂交鹅掌楸的推广应用。自20世纪70年代以来,许多学者对杂交鹅掌楸的组织培养和快繁技术进行了一些探索,但始终没有突破性进展,至今未能形成生产性大规模应用(沈惠娟,1992;陈金慧等,2002)。林木的体细胞胚胎发生研究始于20世纪70年代后期,90年代初得到迅速的发展,并获得了极大的成功。自从1985年挪威云杉体细胞胚胎发生首次报道以来(Gupta et al., 1985),已有40多种木本植物获得了体细胞胚,并且火炬松(Pinus taeda)、挪威云杉(Picea abies)、花旗松(Pseudotsuga menziesii)和辐射松(Pinus radiata)等树种的体细胞胚诱导和植株再生已成功应用于生产实践(施季森,2000)。由于植物体细胞胚具有数量多、一旦形成体细胞胚就能够快速发育成再生植株的特点,以体细胞胚胎发生技术快速繁殖种子来源困难的杂交鹅掌楸,具有重要的理论意义和应用价值。本研究在国家“948 ”项目的资助下,成功利用杂交种体细胞,建立了杂交鹅掌楸体细胞胚胎发生技术和快速成苗体系,开辟了杂交鹅掌楸产业化开发的新途径。
1 材料与方法 1.1 材料来源及处理选用鹅掌楸3个基因型,代号为N, F和X8,于花期用北美鹅掌楸花粉进行人工控制授粉。在南京地区授粉后约8周,开始采样,以寻找最佳取样时间。采下的聚合翅果于4℃条件下低温冷藏,接种时剖开聚合果并除去种翅。去翅材料以洗洁精预处理去表面油污,自来水冲洗30 min,蒸馏水冲洗3次,75%的乙醇处理20 s,0.1%的氯化汞表面灭菌5 min,无菌水冲洗4次。无菌状态下剥去种皮,将胚性材料或胚接种于诱导培养基。
1.2 胚性愈伤组织诱导诱导培养分别以MS,1/2MS培养基为基本培养基,附加激素是浓度分别为1.0, 2.0, 3.0, 4.0, 5.0 mg·L-1的2, 4-D,0.1, 0.2, 0.5 mg·L-1的6-苄胺基嘌呤(6-BA)。另加500~1 000 mg·L-1的天然复合物水解酪蛋白,蔗糖40 g·L-1。杂交鹅掌楸含有较多的单宁类物质,培养过程中培养物容易褐变,在培养基中加入1~5 mg·L-1的维生素C能有效防止褐变发生。培养基以6.5 g·L-1的琼脂粉作固化剂,pH 5.8, 121℃高温高压灭菌16 min。
将试验材料接入诱导培养基,每个培养皿接种4个,分别置光照与黑暗条件下培养,温度22 ~27℃,每20 d更换一次培养基。
1.3 胚性细胞状态调整待培养物长出白色透明愈伤组织后,在无菌条件下将其切开,转至愈伤组织调整培养基。调整培养基中无机盐浓度,改用MS基本培养基,提高蔗糖浓度以改变渗透压,蔗糖浓度分别为40,50,60,70,80 g·L-1。培养基其它成分同诱导培养基。接种物全部转移至光照条件下培养,光照12 h·d-1,光照度为1 200 lx,室温为22~27℃。
1.4 体细胞胚胎发生在状态调整培养基上培养4~6周后,胚性愈伤组织逐渐变得紧密,渐呈颗粒状。转至体胚发生培养基上,采用MS基本培养基,附加的激素组合分别是0.5,1.0, 2.0, 4.0, 5.0 mg·L-1的ZT, 0, 0.1, 0.5, 1.0 mg·L-1的IAA,蔗糖浓度40 g·L-1。22~27℃光照培养。
1.5 体胚发育及植株再生ABA可以促进体细胞胚的成熟和发育过程,将胚性培养物分别在含有0,0.01, 0.05, 0.1, 0.5, 1.0 mg·L-1ABA的培养基中培养,每种浓度各接种3个培养皿,20 d后转入MS培养基使其继续发育,球形胚经过近2个月的发育,会依次经过球形胚、心形胚、鱼雷胚和子叶胚阶段而发育成苗。统计每皿发育正常的幼苗平均数。
2 结果与分析 2.1 亲本基因型与外植体发育状况对体胚发生的影响在参试的3个基因型中(表 1),X8号亲本上采集的聚合果各个发育阶段试材均未能诱导出体细胞胚,而亲本N和亲本F的诱导频率较高,胚性愈伤组织诱导出来以后,多数能顺利产生体细胞胚。
|
|
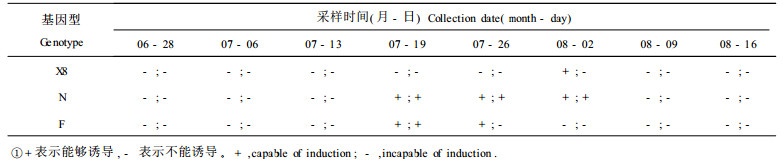
从采样时间看,南京地区7月19日至8月2日期间采集的样品,诱导胚性愈伤组织和体细胞胚的效果都较好。说明采样时间即幼胚发育阶段对体细胞胚胎的诱导起着至关重要的作用,球形期至子叶前期的幼胚对体胚诱导都有效(图版Ⅰ-1)。但是采样时间过早幼胚发育不好或过迟胚胎已经发育成熟,都不利于体胚的诱导。

|
图版Ⅰ Plate Ⅰ |
木本植物胚性愈伤组织的诱导一般在黑暗条件下进行。在试验中发现(表 2),杂交鹅掌楸在黑暗与光照条件下均能诱导胚性愈伤组织产生,但黑暗条件下的诱导频率远远高于光照培养的诱导频率。
|
|
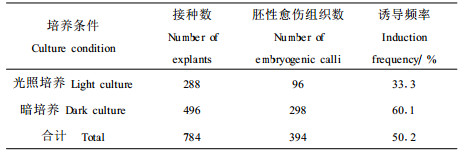
另外,试验发现,培养温度是影响杂交鹅掌楸体胚发生的另外一个关键因素,温度超过30℃,胚性愈伤组织将会永久失去体细胞胚胎发生能力。
2.3 培养基成分与胚性愈伤组织的诱导幼胚接入诱导培养基后约15 d开始启动,长出的愈伤组织晶莹、透明。25 d左右,有些发生少许褐变,经继代后,褐变停止,并开始长出白色、快速增殖的胚性愈伤组织,10 d左右的时间,即可长满整个培养瓶。这种愈伤组织在解剖镜下观察,发现细胞多为圆球状,水分含量多,细胞质稀薄,结构较松散。
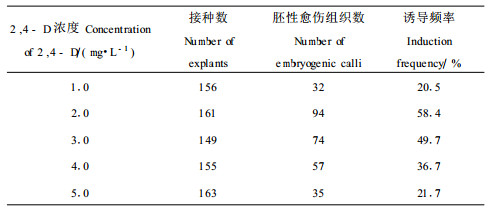
培养基中外源激素的配比对胚性愈伤组织的诱导及生长有很大的影响,特别是2, 4-D的浓度起着至关重要的作用(表 3)。虽然在各种2, 4-D浓度的培养基上都能诱导出胚性愈伤组织,但诱导率存在很大差异。当2, 4-D浓度为1.0 mg·L-1时,诱导率低,愈伤组织生长较慢,一个月以后会有褐变现象产生。当2, 4-D浓度为5.0 mg·L-1时,诱导率也较低,愈伤组织生长迅猛,到了细胞状态调整阶段以后,由于生长素的后续效应,细胞多为胞质稀薄的薄壁细胞,不利于体细胞胚胎发生。当2, 4-D浓度分别为2.0,3.0 mg·L-1时,胚性愈伤组织诱导频率较高,细胞生长状态容易调整,体细胞胚胎发生相对比较容易。
|
|
试验证明,渗透压对体细胞胚胎的形成起着重要的作用。在裸子植物的体细胞胚胎发生研究中,通常利用肌醇(黄健秋等,1995)或聚乙二醇(Li et al., 1997)作渗透剂,提高培养基的渗透压。在研究中发现,利用蔗糖作渗透剂,对于胚性细胞的伤害较小。随着渗透压的提高,愈伤组织含水量逐渐降低而变得干燥、紧密,培养一个月后,会形成颗粒状的愈伤组织,进而发育成体细胞胚(图版Ⅰ-2)。说明随着渗透压的提高,体胚诱导能力增强。但是渗透压过高,超出一定范围,其诱导体细胞胚胎的能力反而下降,在本研究中,蔗糖浓度60 g·L-1时的渗透压为最适渗透压。
另外值得一提的是,及时将胚性愈伤组织转移至光照条件下培养非常重要,因为光照培养条件下,能有效控制细胞的分裂速度。
2.5 细胞分裂素在体细胞胚胎发生中的作用经蔗糖渗透调节后的胚性细胞,细胞质逐渐变得浓厚,细胞新陈代谢旺盛,细胞中沉积大量的淀粉粒,为体细胞胚胎发生做准备。当培养细胞接种到以细胞分裂素为主的体胚发生培养基上,培养基中高浓度的细胞分裂素会缓和愈伤组织诱导阶段生长素的后续效应,改善细胞状态,经过一段时间的培养,来源于胚性愈伤组织表面的胚性细胞便会分化成为球形胚(图版Ⅰ-3,4)。
细胞分裂素ZT是鹅掌楸体细胞胚胎发生过程中的关键因素。随ZT浓度升高,体细胞胚胎发生频率亦随之上升,但浓度过高也会有副作用产生,即细胞中容易沉积叶绿素,导致其老化死亡。ZT浓度为1.0, 2.0, 4.0 mg·L-1时,都能诱导体细胞胚胎发生。
2.6 体细胞胚的发育和成熟体细胞胚胎发生研究中最大的困难就是畸形胚的发生。在有些裸子植物的研究中,培养基中加入适当浓度的ABA,胚发育不正常的情况,如子叶合生、早熟发芽等会受到抑制(唐巍等,1996)。在本研究中发现(表 4),加入适量的ABA(0.05 mg·L-1),不仅能够抑制畸形胚的产生(图版Ⅰ-5),而且能够提高体细胞胚的萌发率。
|
|
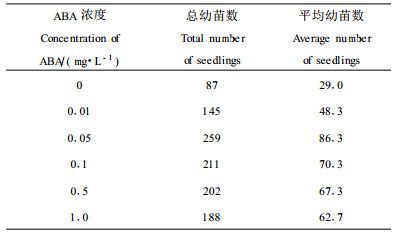
研究表明,一旦球形胚形成以后,立即转入不含任何激素的MS培养基,也能有效减少畸形胚的发生。并且体细胞胚具有两极性,比较容易形成发达的根系,有效解决了鹅掌楸常规组织培养中丛芽不生根或生根难的问题(图版Ⅰ-6~8)。
3 结论与展望研究证明,体细胞胚胎发生能力与外植体的发育状态有着极为显著的联系,而胚胎的发育与当年积温有关。所以在采集外植体时,应该根据当年积温,选取球形期至子叶前期材料为好。
在植物材料发育的不同阶段,有着不同的营养需求,需要不同的基本培养基,这可能与氮元素、钙离子等的吸收有关。本试验在愈伤组织诱导阶段,使用1/2MS培养基,而到了胚性组织的诱导阶段,提高无机盐的用量,改用MS基本培养基,一方面是因为在此阶段,培养物的营养需求旺盛,另一方面,也可以提高培养基的渗透压,利于体细胞胚胎发生。
本试验建立的杂交鹅掌楸体细胞胚胎发生体系,不仅繁殖系数高,再生植株移栽成活率高,而且稳定性好,已经连续3 a获得了再生植株。近3万株再生植株的田间试验结果表明,再生植株保持了明显的杂种优势,并在生长势、株型结构上明显优于同期扦插苗。
进一步研究利用生物反应器大规模生产杂交鹅掌楸体细胞胚,这样可以节约生产成本,提高生产效率;由于体胚发生体系较为稳定,可以在分子水平上研究胚胎发生的机理;体细胞胚胎发生系统是遗传转化研究的良好受体系统, 利用农杆菌介导的转化方法,可以转入有用抗性基因,进行遗传改良研究。
陈金慧, 施季森, 诸葛强. 2002. 杂交鹅掌楸的不定芽诱导及植株再生. 植物生理学通讯, 38(5): 459. |
黄健秋, 卫志明. 1995. 针叶树体细胞胚胎发生的研究进展. 植物生理学通讯, 31(2): 85-90. |
季孔庶, 王章荣. 2001. 鹅掌楸属植物研究进展及其繁育策略. 世界林业研究, 14(2): 8-14. DOI:10.3969/j.issn.1001-4241.2001.02.002 |
李周歧, 王章荣. 2001. 鹅掌楸属种间杂种苗期生长性状的遗传变异与优良遗传型选择. 西北林学院学报, 16(2): 5-9. DOI:10.3969/j.issn.1001-7461.2001.02.002 |
南京林产工业学院林学系育种组. 1973. 亚美杂种马褂木的育成. 林业科技通讯, (12): 10-11. |
沈惠娟. 1992. 木本植物组织培养技术. 北京: 中国农业科技出版社, 58-67.
|
施季森. 2000. 迎接21世纪现代林木生物技术育种的挑战. 南京林业大学学报, 24(1): 1-6. DOI:10.3969/j.issn.1000-2006.2000.01.002 |
唐巍, 杨映根, 桂耀林, 等. 1996. 松柏类植物体细胞胚胎发生的研究与应用. 植物学通报, 13(1): 25-31. |
Gupta P K, Durzan D J. 1986. Plantlet regeneration via somatic embryogenesis from subcultured callus of mature embryos of Picea abies(Norway spruce). In Vitro Cellular and Developmental Biology-Plant, 22: 685-688. DOI:10.1007/BF02623484 |
Hakman I, von Arnold S. 1985. Plantlet regeneration through somatic embryogenesis in Picea abies(Norway spruce). J Plant Physiol, 121: 149-158. DOI:10.1016/S0176-1617(85)80040-7 |
Li X Y, Huang F H, Gbur E E. 1997. Polyethylene glycol-promoted development of somatic embryos in loblolly pine (Pinus taeda L.). In Vitro Cellular and Developmental Biology-Plant, 33(3): 184-189. DOI:10.1007/s11627-997-0019-5 |
Merkle S A, Sommer H E. 1986. Somatic embryogenesis in tissue culture of L. tulipifera. Can J For Res, 16: 420-422. DOI:10.1139/x86-077 |