 2003, Vol. 39
2003, Vol. 39文章信息
- 孙鹏森, 刘世荣.
- Sun Pengsen, Liu Shirong.
- 鄂尔多斯地区两种典型沙质荒漠化阶段微气象特征的比较研究
- COMPARISON OF MICRO-METEOROLOGICAL CHARACTERISTICS OF DIFFERENT STAGE IN DESERTIFICATION AREA ERDUOSI
- 林业科学, 2003, 39(4): 9-15.
- Scientia Silvae Sinicae, 2003, 39(4): 9-15.
-
文章历史
- 收稿日期:2002-03-11
-
作者相关文章


沙质荒漠化是在自然、生物和人为因素等多种驱动力交互作用下产生的土壤沙化及生产力和土地退化的过程,是生物地球物理过程和生物地球化学循环产生的物流、能流过程变化所形成、演变的结果。因此,研究沙质荒漠化过程中的生态学过程,特别是水分及能量的转化与平衡规律对揭示沙质荒漠化的形成、演变机制和植被对沙质荒漠化过程的响应至关重要,这是沙质荒漠化过程基础理论研究的核心内容。Hobbs(1994)、Bryant (1990)和Sala (1997)曾就热带干草原土壤水分的垂直分布和温带草原植物功能类型进行过研究,认为土壤水分梯度和持久性决定了植被演替的方向,草本趋向于利用土壤上层0~50 cm的水分,而多年生木本植物则较多利用下层水分。Philllips(1993)曾就这一现象提出假设:旱地的土壤水分直接与植被覆盖度、降雨量和水蚀成正相关,而与反射率、温度和风蚀成负相关。国内相关研究很少,李胜功(1999)认为:沙化地区的微气象特征中反射率是区别不同沙漠化土地发展进程的重要指标,随着沙漠化的发展,净辐射和潜热交换趋于减少,风速分布有偏离分布的趋势。
1 研究地区概况研究地点位于中国科学院植物研究所鄂尔多斯沙地草地生态研究站及其周围地区,东经109 °51′,北纬39°02′。海拔1 355 m,地处内蒙古鄂尔多斯高原毛乌素沙地北缘伊金霍洛旗霍洛苏木石灰油大队。伊金霍洛旗地处鄂尔多斯高原特殊的地理景观多层次生态过渡带。属暖温带干旱、半干旱大陆性季风性气候、年均降水量353.6 mm,年均蒸发量2 388.7 mm,其特殊的环境条件适合于灌木和半灌木的生长和发育。在所有的灌木、半灌木中,尤其以半灌木油蒿(Artemisia ordosica)最占优势,其分布面积占鄂尔多斯高原总面积的47.3%,为鄂尔多斯高原植被的优势种和建群种,正向演替可形成地带性植被本氏针茅(Stipa bungenana)群落。但是,经过度放牧破坏后,退化为油蒿+牛心朴子(Cynanchum komarocii)群落和牛心朴子群落(王庆锁等, 1997)。
为阐明沙质荒漠化过程的演化机理,在鄂尔多斯地区选取沙质荒漠化过程中的两个典型阶段——油蒿群落和牛心朴子群落进行了连续的定位观测。在沙质荒漠化类型上,两者都属于风蚀沙质荒漠化,土地类型分别属于硬梁地草场和软梁地草场,油蒿群落的植被盖度达60%~70%,牛心朴子仅为20%~30%,牛心朴子的出现是该地区沙质荒漠化程度由中度向重度转化的重要标志,因此了解两者的水分与能量特征的变化对于分析沙质荒漠化的驱动与成因有着非常重要的意义。
2 研究方法分别在典型的油蒿和牛心朴子群落样地内安装全自动气象站,进行不同沙质荒漠化阶段的对比观测研究。群落斑块面积不小于1 km2, 两站相距1.5 km左右。自动气象站(Compbell Scientific, Inc.),配备CR10X数据采集装置。微气象因子梯度观测的配置设定如下:大气降雨量P、太阳总辐射Rt,净辐射Rn,空气温度Ta, 相对湿度RH,分2m高处和4m高处,风速Vw, 风向Wd;土壤温度St和湿度Sm的测定深度分别为:10cm, 20cm, 40cm, 80cm。利用自动气象站配置程序,设定采样频率,所有观测因子10min同步采样1次,每隔30min作1次平均值,存入系统。观测时间为2000年5月—2001年9月。
数据分析采用SPSS 10.0统计分析软件包。
3 结果与讨论 3.1 油蒿群落和牛心朴子群落的辐射收支状况下垫面所接收的净辐射能Rn=LE+H+G,其中LE为潜热,H为显热交换,G为土壤热通量。其中显热交换项与潜热项的比值(波文比,β=H/LE),是利用微气象法求算群体水平蒸散的一个很重要的参数,也是衡量局域小气候的一个综合指标。另外,根据辐射平衡式:Rn=Rt-Rr-I(Rr反辐射,I为净长波辐射,即地表向上长波辐射减去大气向下长波辐射),反射率α(α=Rr/Rt)则由下垫面的性质直接影响,因此,在相同的太阳总辐射条件下,下垫面不同,所接收的Rn值不同。研究区夏季植物生长旺盛,形成一定的地表覆盖率,因此,反射率降低,净辐射所占比例较大,约占太阳总辐射的57.6%,冬季,随着太阳总辐射的降低,净辐射也随着降低,但是由于植被枯萎,地表覆盖率降低,反射率α增高,所以,净辐射的比率降低至43.2%,从全年平均值看,净辐射占总辐射的56.5%。净辐射最高值在7月,可达1 015 W·m-2,最低12月,为363 W·m-2 (图 1-a)。研究区油蒿群落具有相对较好的盖度(60%~70%),反射率低,Rn值较高;而牛心朴子群落高度退化,植被盖度低(仅为20%~30%),裸露的沙面对太阳的反射较强,因而下垫面所接收的Rn值较低。(图 1-a)为两种下垫面净辐射Rn的比较,油蒿群落比牛心朴子群落吸收更高的净辐射能,特别是在辐射强度较高时,二者的差距更大。生态系统的能量流动首先应当从其接收的净辐射能开始,因此我们可以认为,沙质荒漠化过程的产生,首先是由于改变了生态系统中下垫面的性质,因而改变了系统中的能量分配,而能量的分配与生态系统中最重要的物质循环—水分的循环密切关联的。将二者净辐射的差值进行研究,可以明显看出以下规律:1)10月到第二年5月份,两者差别较小(±100 W·m-2以内),但油蒿的Rn减去牛心朴子的Rn基本为正值;5~9月份,二者差别较大(±200 W·m-2变动),虽然出现个别负值,但仍以正值为主(图 1-b)。

|
图 1 油蒿群落和牛心朴子群落的所接受的太阳净辐射比较(2000~2001) Fig. 1 Comparison of net solar radiation(Rn) in A. ordosica community C.komarocii community (2000~2001) a: 月净辐射最大值 Monthly maximum Rn ——油蒿 A. ordosica ---牛心朴子 C. komarocii b: 月平均净辐射之差 Difference of monthly average Rn |
根据两台自动气象站在2000年5月26日至2001年9月22日的测定记录,该区降雨总量平均为14 5.9 mm,两站的降雨量观测并无显著差异(表 2)。图 4是每日降雨的累计值,可以看出,该区降雨最大月份出现在7、8两个月,总降雨为全年的66%,最高8月份累计降雨量43.2 mm,低的不及1.0 mm。4月份观测到的降水量达13.3 mm,可能是由于雨量筒中的雪融化所致,这些水分应当包括12月到来年3月期间的降雪融化水分(图 3)。
|
|

|
图 4 不同深度月平均土壤温度的季节变化规律(2000—2001) Fig. 4 Vertical Change of soil temperature in different communities: A. ordosica and C. komarocii community (2000—2001) a:油蒿群落A. ordosica b:牛心朴子群落C. komarocii |

|
图 3 月降雨在两年内的分配(2000~2001) Fig. 3 Monthly precipitation graph in the study area during 2000~2001 |
土壤含水量的垂直梯度非常明显,就总体而言,牛心朴子群落的土壤含水量在80 cm深度维持在0.08 g·g-1的水平,低于其他各层,土壤含水量的大小次序是20cm>10cm> 40cm>80cm;油蒿80cm的土壤含水量明显高于其他各层,其次为10cm, 20cm和40 cm。影响土壤含水量因子较多,植被因素只是其中之一,由于所选择的研究地点相距较近,可以基本排除降雨量的影响(经检验,降雨无差异,表 2)。因此,植被和土壤的物理结构是影响含水量的重要因子。经过野外初步调查发现,油蒿群落主要分布在固定和半固定沙地,牛心朴子基本分布在半流动沙地,其土壤的蓄水作用明显不同,油蒿群落的土壤含水量略高于牛心朴子。另外,由于两个群落分别处于不同的梁地上,下层水分含量差别也很大,油蒿的硬梁地深层具有一定的保水作用,降雨后水分不易透过,因此,80cm处的土壤水分明显高于其他各层。而牛心朴子的软梁地沙层较厚,降水后水分迅速入渗,而表层由于蒸发作用迅速干化,因此总体来说土壤含水量较低。
从土壤水分稳定性来看,越往深层,土壤含水量越稳定,土壤表层,尤其是0~20 cm之间的土层土壤水分变动幅度大。首先,降水是影响表层土壤水分的关键因素,从2000年5月开始观测至2001年9月,最大的一次降雨过程发生在2001年8月,月降雨总量达45.2 mm,各层土壤水分均发生不同程度的上升。而其它时间,月降水都低于25 mm,土壤水分只在40 cm的深度发生变化(图 2)。另外,融雪也会对土壤含水量有某种程度的影响。从2月中旬到4月初,10 cm、20 cm土层含水量有缓慢回升的趋势,但此时并没有降雨(图 3)。10月中旬到翌年2月中旬,土壤含水量基本保持平稳,而且各层相差不大。因此可以得出这样的结论,如果一场降雨低于45 mm,则基本上只能影响到0~40 cm土壤的含水量,而且持续时间较短,但对80 cm的深层几乎没有影响;而当月累计降雨超过45 mm,80 cm深层的土壤含水量也会大幅度提高,并且持续时间较长。早春的融雪可以使20 cm以内的土壤水分提高0.02~0.04 g·g -1。这一点非常重要,沙质荒漠化地区的降水普遍偏少,但对土壤水分的有效层次及降水作用深度决定了沙漠中植被的分布(Sala,1997)。在这一点上两群落表现的规律基本一致,只是在基本相同的降水条件下,牛心朴子表层土壤水分波动明显大于油蒿,主要原因是油蒿地上生物量和盖度小,降雨时植物层截留蒸发少,全部水分都补充到表层土壤中,所以土壤水分上升很快;而晴天蒸发又快,所以水分下降也快。另外两者的根系分布深度对土壤水分含量也有影响。牛心朴子根系的范围明显浅于油蒿(表 1),很难发挥对深层土壤水分的调蓄作用。

|
图 2 不同深度土壤含水量的季节动态(2000—2001) Fig. 2 Seasonal variation of soil moisture in different depths of two communities(2000—2001) a:油蒿群落A. ordosica b:牛心朴子群落C. komarocii |
|
|
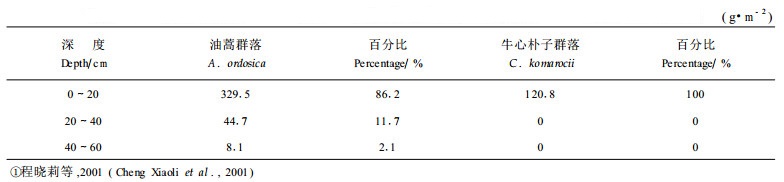
尽管观测时段正值欠水年,但总的说来,该区土壤水分并不低,尤其是深层土壤水分。从油蒿群落的土壤水分图可以看出,植物生长可利用的水分都在深层,而这样的深度是许多现存的灌木和草本根系达不到的,根据调查,油蒿群落最深根系可达60 cm,而牛心朴子群落根系全部分布在0~20 cm的深度(表 1)。因此,油蒿群落能够充分利用大部分降水输入的水分,而牛心朴子群落却只能利用较小雨量的降水。另外,牛心朴子群落盖度低,大部分净辐射用于土壤水分的蒸发,可能是引起深层土壤含水量降低的原因之一。
3.3 土壤温度土壤温度的时间序列规律除了日周期外,还有明显的年变化周期。从测定之日(5月26日)起到7月23日,虽然还有个别的降温天气,土温也呈现相应的波动,但总的来说,各层的温度是呈现上升趋势的,表层(10 cm)月平均土温最高可达27.7℃。7月以后,各层温度逐渐下降,一直到来年1月上层(10 cm)的月均土温已经降到最低点-6.4℃左右,之后又开始回升。其他各层大致规律也是如此,只是峰值大小及出现的时间有明显的差别,从时间上看,10、20、40、80 cm日平均土温最高值出现的时间依次是:7月21日、7月21日、7月22日、7月25日;大小分别为:32.9℃、31.6℃、28.5℃、23.7℃。日平均土温最低值出现的时间依次是1月28日、1月28日、1月29日、1月31日;大小分别为:-8.7℃、-7.9℃、-5.1℃、-1.6℃。可见,深层土温的峰值变化要比表层晚1~2 d,高温差值为9.2 ℃,低温为7.1℃。从各层比较来看,越往表层,土壤温度的变化越剧烈,越往内层,越趋于稳定,这在图 4中可以很明显的看出来。注意两个重要的转折点:9月26日以后,表层土温开始低于深层,虽然还有个别温度回升的天气,但总的说来温度高低的次序已经完全逆转。从第2年3月17日开始,地温又逐渐从表层开始升高(图 4)。
牛心朴子和油蒿日均最低温度相比,以牛心朴子减去油蒿来看,10、20、40、80 cm差值依次为-0.22℃、-0.18℃、0.16℃、0.44℃;日均最高温度,同样以牛心朴子减去油蒿来看,10、20、40、80 cm差值依次为0.88℃、1.34℃、0.75℃、-0.09℃。这些数据表明,油蒿群落更有利于维持地温的恒定。从地温极大值和极小值出现的时间来看,两者是完全一致的。
3.4 风速通过对近地面风速(地上2 m)的精确测定可以看出:两群落的风速略有差异,差值变动在0.2~0.5 m·s-1之间,牛心朴子高于油蒿群落,10月至翌年4月份差值明显,而5~9月的生长季差别相对较小。这说明,油蒿对近地面风速的削减作用大于牛心朴子群落,并且这一作用在秋冬季节略大于生长季(图 5)。

|
图 5 近地面层(地上2 m)平均风速的季节规律(2000—2001) Fig. 5 Average wind speed of A. ordosica and C. komarocii community, 2 meters above ground(2000—2001) |
风是该区沙质荒漠化的最重要的成因,因而,削减风速能够大大减轻沙质荒漠化的进程。而近地面层风速受下垫面的性质影响很大,即下垫面粗糙度的影响,在相似的地貌类型下,地表植被是影响粗糙度的主要因子。粗糙度可以通过植被高度近似计算(Z≈0.1 H,H为植被高度)。因此,植被高度与下垫面的粗糙度呈正比的关系。根据调查,油蒿群落地上部分的生物量可以分布到60 cm,而牛心朴子的生物量则主要集中在30 cm(程小莉,2001)。
自零位移高度起,风廓线一般为对数分布(翁笃鸣,1981),通过选取不同季节典型天气的风速2 m和4 m进行比较,就很容易看出不同下垫面粗糙度对风速的影响。牛心朴子地上2 m和4 m之间的风速差别非常小,通常小于0.1 m·s-1,而油蒿群落的风速差异则很明显,削减风速在0.2~0.5m·s-1之间,并且这一规律在不同季节都表现一致,即在各季节,油蒿群落都有着比牛心朴子群落更强的削减近地层风速的作用,从而更有效的防止了风蚀沙质荒漠化的发生(图 6 a~h)。

|
图 6 油蒿(a~d)和牛心朴子群落(e~h)不同高度风速的比较 Fig. 6 Comparison of wind profile in A. ordosica (a~d) and C. komarovii (e~h) |
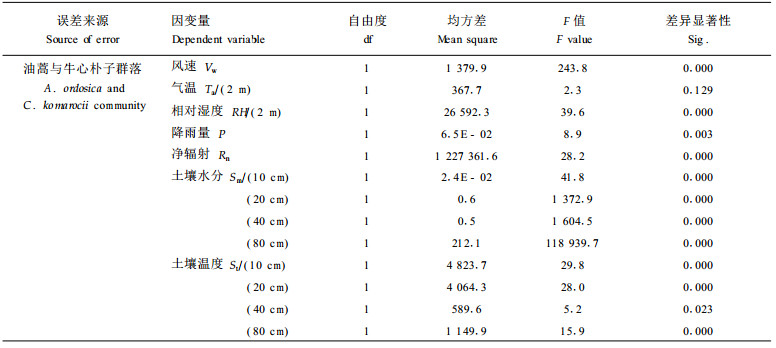
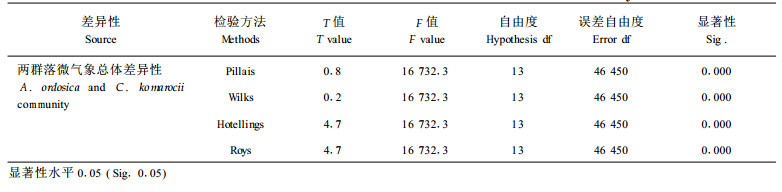
以上分析分别从不同角度阐述了不同沙质荒漠化阶段,即油蒿群落阶段和牛心朴子群落阶段不同微气象因子的差异,而多因变量线性方差分析(MANOVA)的检验结果表明,两群落除气温、降雨量、40 cm土温不存在显著差异(0.001水平)外,其余各因子包括净辐射、风速、相对湿度、土壤各层水分及10 cm、20 cm、80 cm土壤含水量均存在明显差异(表 2)。MANOVA分析表明,四种检验方法下两群落气象因子总体上存在显著性差异(表 3)。
|
|
通过对不同沙质荒漠化阶段的油蒿群落和牛心朴子群落的总辐射、净辐射、土温、土壤含水量、风速、降雨量等微气象因子的分析,可以得出以下结论:
油蒿群落比牛心朴子群落吸收更高的净辐射能,特别是在辐射强度较高时,二者的差距更大。
总体上看,油蒿群落的土壤含水量略高于牛心朴子。在基本相同的降水条件下,牛心朴子表层土壤水分波动明显大于油蒿。牛心朴子根系的范围明显浅于油蒿,很难发挥对深层土壤水分的调蓄作用。
从各层土壤温度的变动规律来分析,油蒿群落更有利于维持地温的恒定。
油蒿对近地面风速的削减作用大于牛心朴子群落,并且这一作用在秋冬季节略大于生长季,从而某种程度上防止了风蚀沙漠化的发生。
MANOVA分析表明,两群落气象因子总体上存在显著性差异,而气温、降雨量、40 cm土温不存在显著差异。
草地沙质荒漠化的过程,总是伴随着生态系统能量和物质运动的变化,而这种变化,在很大程度上通过生态系统的微气象特征显现出来。本文分析的几个特征是其中变化比较明显的。而由这些气象特征的变化所引起的群落水分蒸散的变化、地下水的变动等,以及由此引发的大尺度上植被格局的演变形式,是需要进一步探明的重要课题。
程小莉, 安树青, 陈兴龙, 等. 2001. 鄂尔多斯草地荒漠化过程与植被生物量变迁的关系. 林业科学, 37(2): 13-20. |
李胜功, 何宗颖, 申建友. 1994. 奈曼沙漠化对草地微气象特征影响的研究. 大气科学, 18(6): 758-763. DOI:10.3878/j.issn.1006-9895.1994.06.13 |
李胜功, 赵哈林, 何宗颖, 等. 1999. 不同放牧压力下草地微气象的变化与草地荒漠化的发生. 生态学报, 19(5): 697-704. DOI:10.3321/j.issn:1000-0933.1999.05.018 |
林业部科技司编.森林生态系统定位研究方法.北京: 中国科学技术出版社, 1994: 246-259
|
王庆锁, 梁艳英. 1997. 油蒿群落植物多样性动态. 中国沙漠, 17(2): 159-163. |
翁笃鸣, 陈万隆著.小气候和农田小气候.北京: 农业出版社, 1981
|
Bryant N A. 1990. Measuring the effect of overgrazing in the Sonoran Desert. Climate Change, 17: 243-264. DOI:10.1007/BF00138370 |
Hobbs T J. 1994. A model of soil moisture balance and herbage growth in the arid rangelands of central Australia. Journal of Arid Environment, 28: 281-298. DOI:10.1016/S0140-1963(05)80049-7 |
Philllips J D. 1993. Biophysical feedbacks and risks of desertification. Annals of the Association of American Geographers, 83: 630-640. DOI:10.1111/j.1467-8306.1993.tb01957.x |
Sala O E. Plant functional types in temperate semi-arid regions. In: Smith. eds. Plant Functional Types. Cambridge University of Press, 1997
|