 2003, Vol. 39
2003, Vol. 39文章信息
- 李明鹤, 沈宝仙, 李传涵, 梅勇, 严明学.
- Li Minghe, Shen Baoxian, Li Chuanhan, Mei Yong, Yan Mingxue.
- 杉木无性系选择与木材生产相结合的研究
- CLONAL SELECTION IN COMBINATION WITH TIMBER PRODUCTION OF CHINESE FIR (CUNNINGHAMIA LANCEOLATA)
- 林业科学, 2003, 39(3): 157-162.
- Scientia Silvae Sinicae, 2003, 39(3): 157-162.
-
文章历史
- 收稿日期:1999-11-09
-
作者相关文章


2. 湖北省虎爪山林场 京山 431815
2. Tiger Paw Forest Farm, Hubei Province Jingshan 431815
自1964年以来, 我国建立了大面积的杉木种子园, 选出了许多优良家系和优良种源。但是, 种子产量不稳定, 控制授粉成本高, 育苗技术复杂, 是实生苗造林中存在的问题。无性系林业可以获得更高的增产效果,凡可以低成本大规模无性繁殖的树种, 均应采用无性系林业(朱之悌, 1986)。10余年来, 扦插育苗和成年树复壮技术有了重大进步(李明鹤等, 1990a; 1990b; 1990c), 这为实现完整的无性系林业(full clonal forestry)铺平了道路。但是, 传统的无性系育种法所需时间长, 遗传基础窄, 不适合于杉木。1986年, 李明鹤提出了由种子繁殖过渡到无性系林业的无性系选择方案。它分为两步:第1步, 以“大群体”为基础, 用优良群体的实生苗建立采穗圃, 用扦插苗代替实生苗造林;第2步, 结合造林进行简单的无性系测验, 在采穗圃中淘汰不良的无性系, 并用保留的无性系造林。上述过程每隔几年重复一次。这种方法既能满足当前造林对大量苗木的需要, 节省时间, 简单、经济, 没有风险, 也可选出优良的无性系品种来(李明鹤, 1993; Li Minghe et al., 1999c)。试验证明, 该方案第1步是可行的(李明鹤等, 1990a; 胡伯智等, 1999; Li Minghe et al., 1999b)。本文将用新的试验对第2步的可行性进行论证, 并根据试验结果对杉木无性系选择策略进行了讨论。
1 材料和方法 1.1 无性系的来源穗条采自湖南省通道县林科所采穗圃, 母本系会同种子园自由授粉实生苗。1991年春选择生长正常, 无病虫害, 萌条多的母本共1 000株, 从每母本上各采条15根, 扦插为1行, 每行为1个无性系, 共1 000个无性系。1992年春, 根据成苗率、苗木生长、抗性等情况, 用目测法选优, 保留552个无性系, 并分为两群,其中一群含252个无性系引入湖北省京山县虎爪山林场营造无性系测验林。
1.2 田间试验设计每无性系用扦插苗6株。其中1株用于建立采穗圃; 另5株用于营造无性系测验林,随机完全区组设计, 单株小区, 5次重复, 分设在2个地点。区组为长方形, 252株苗木沿等高线方向栽成12行, 21列; 株行距1.3 m×2.0 m, 5个区组共栽无性系苗1 260株。各区组分散在用当地1 a生实生苗营造的人工林中。人工林株行距2 m×2 m。区组四周各划出4行总面积与区组相等的人工林为对照。
1994和1997年底, 根据无性系的表现, 按15%的比率对无性系林各进行1次去劣间伐, 分别伐去38个和32个, 共70个无性系。
1.3 区组内土壤差异的校正为比较各无性系的相对优劣, 区组内的土壤差异根据间比法的原理用等级差距法从2个方向进行校正。结果表明, 由于区组的面积不大, 同一区组上、下、左、右各4个对照行或列的平均值相差很小, 表明区组内土壤差异不大。但是, 无性系比较时, 所有的数值均按此方法进行了校正。
1.4 材积的计算每年冬季测量试验林的树高和直径。从1995年开始用下列公式计算立木材积:V=(π/4)D1.32(H+3)0.41,式中,V为材积;D1.3为胸径;H为树高。
1995年以前用下列公式计算立木材积:V=(π/4)D02(H+3)0.41×0.482,式中,D0为地径, 0.482为根据1995年胸径与地径求得的换算系数。1995年用2种公式计算所得材积相等。
1.5 定植苗大小的校正1 a生扦插苗通常比同龄实生苗高8~10 cm, 直径几乎大1倍。为了得到更正确的结果, 在分析时, 用协方差法将定植时无性系苗高、直径和材积按对照苗高度进行了校正。
1.6 资料整理和分析用微型计算机SAS软件进行统计分析。以单株平均数为单位, 用成对样本T检验法比较无性系和对照的优劣, 用GLM法进行方差分析。
2 结果与分析 2.1 无性系群体与对照的比较会同种子在京山县是优良种源。由于对照设在区组四周, 无性系与对照的平均值巳消除了土壤差异的影响, 因此可直接将各区组历年树高、地径或胸径、材积平均值进行比较。差异显著性测验所得结果列于表 1。可以看出, 自1992年春定植时起至1997年底, 无性系群体树高、直径和材积均显著或极显著地超过了对照。这表明, 在林业生产中, 以大群体为基础, 把已知优良群体进行无性繁殖, 并按无性系的表现定期进行去劣间伐, 则无性系林具有稳定而可靠的增产效果。去劣间伐也在采穗圃中进行, 因此以后营造的人工林可以获得相似的增产效果。
|
|
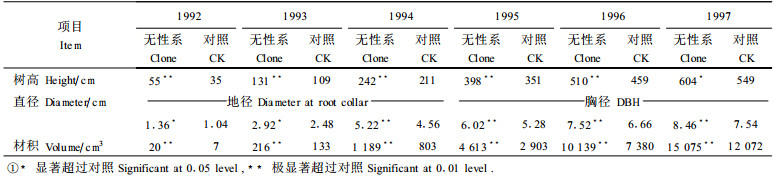
不同年份以及去劣前后树高、直径和材积增长超过对照的绝对值和相对值如表 2。表 2的数字表明, 无性系群体树高、直径和材积超过对照的绝对值随着年龄的增长而增长。例如, 从1992年到1997年, 树高从20 cm提高到66 cm, 直径从0.32 cm到0.96 cm, 材积从13 cm3到4 001 cm3。这表明了无性系群体在遗传上的优势。但是, 增产的相对值则是随着年龄的增长而迅速降低的。例如, 从1992年到1997年, 树高从57.1%降到12.0%, 直径从30.8%降到12.7%;材积从185.7%降到33.1%。如果不是由于1994年和1997年2次去劣曾大幅度地提高了增长率, 其下降的程度还要大, 即, 树高应降至7.2%, 直径降至11.3%, 材积降至11.5%。这预示着, 尽管无性系群体在遗传上有优势, 但由于树高、直径和材积累计值增长得更快, 早期显示的高增产率到25 a或30 a后采伐时将变得非常小。
|
|
从表 2还可以看出, 1994年和1997年所进行的去劣选择明显地提高了遗传增益的绝对值和相对增产率。例如, 1994年去劣后, 材积绝对增值从386 cm3提高到494 cm3, 相对增值从48.1%提高到61.5%;1997年相应地从3 003 cm3提高到4 001 cm3, 相对值从24.9%提高到33.1%, 等等。去劣缓和了增产率随年龄增长下降的趋势。
图 1表示1997年按15%的比例淘汰32个最差无性系的情形。从左至右, 3条曲线分别代表对照、去劣前和去劣后无性系混合体。X1、X2和X3代表它们的平均数。去劣前无性系曲线左侧较陡是由于1994年己进行过一次去劣。由于1997年的去劣间伐, 无性系群体平均数从X2移向X3。淘汰的比率越高, 则增产率提高的数字也越大。但是, 去劣的比率不可过大, 以便为以后的选择留下足够机会。

|
图 1 1997年去劣效果 Fig. 1 Effects of the clonal plantation by roguing in 1997 |
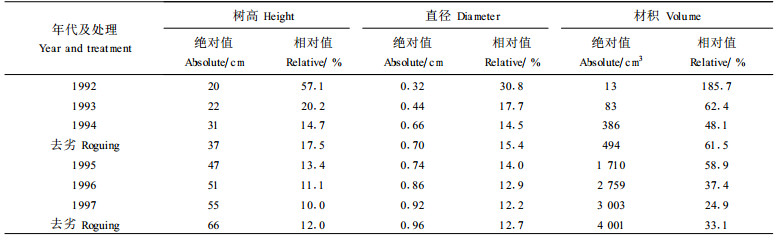
1 a生扦插苗通常比实生苗高8~10 cm, 直径几乎大1倍。本试验所用无性系来源于会同种子园种子所育实生苗, 对照为本地实生苗。会同种子在京山县系优良种源, 因此, 本试验中无性系林树高、直径和材积在定植时及以后超过对照的数量, 应分解为由于较大的苗木和由于种源优势两部分, 并可通过协方差分析进行分解。树高、直径和材积具有相似的变化规律, 因此, 表 3仅列出了材积增产的绝对值及其相对增产率协方差分解结果。表 3重现了表 2中的趋势:材积增产的绝对值随年龄的增长而增加, 但其相对增产率则逐年下降, 而去劣间伐可以提高增产的绝对值与相对值。但是, 值得注意的是, 去劣间伐对种源和定植苗大小有着不同的效果。去劣提高了无性系群体因种源而引起的绝对值和相对值, 如本例中绝对值从1994年的331 cm3提高到458 cm3, 相对值从41.2%提高到57.0%; 1997年相应地从2 564 cm3提高到3 926 cm3和从21.3%到32.5%。但去劣却显著地降低了因苗木大小而引起的影响,如本例中材积增产绝对值从1994年的55 cm3降为36 cm3, 相对值从6.9%降为4.5%; 1997年相应地从439 cm3降至75 cm3和从3.6%降至0.6%。因苗木大小而引起的材积增产率在定植后第2 a为114.3%, 至第7 a时其影响仅为0.6%。这表明, 决定林分后期增产效果的主要因素是苗木的遗传品质, 苗木大小对长期的研究或生产并无太大的重要性。
|
|
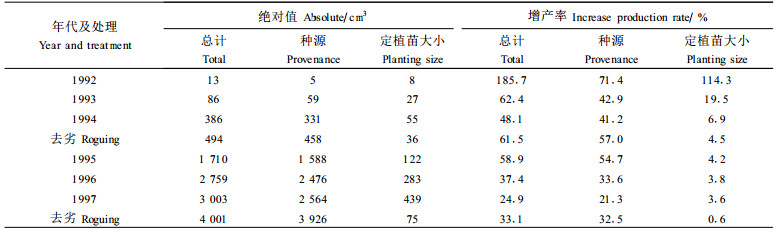
1992、1994和1997年树高、直径和材积方差分析所得F值和差异性概率P值如表 4。表 4表明, 各年区组间F值均很大, 表明各区组被安排在很不相同的环境中。去劣前, 除1997年树高为显著外, 各年无性系间在树高、直径和材积上均有极显著的差异。计算表明, 1994年时, 有的无性系材积超过对照2.62倍, 有的低于对照68%;1997年, 有的超过对照2.72倍, 有的低于对照33%。这从图 1和图 2也可大致看出。去劣使F值减小, 但1997年去劣后直径和材积仍保留了极显著差异, 而且预期还会随着年龄的增长而增加。这说明, 群体在将来有充分的进行去劣选择的潜力。
|
|

|
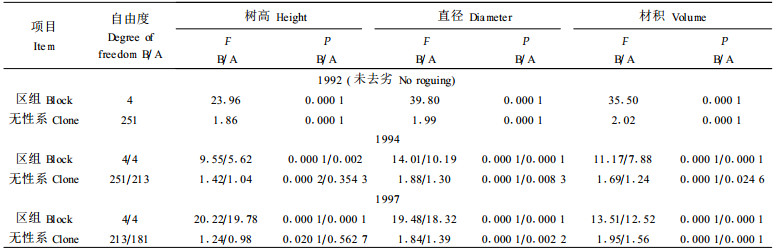
图 2 在1994年选择少数无性系时对1997年材积的影响 Fig. 2 Effect of selecting few clones in 1994 on volume gain in 1997 图中每一个点代表一个无性系, 共214个无性系。横向和纵向虚线分别代表 1994年和1997年按材积选择最好的10个无性系的情形。 Each point stands for a clone and a total of 214 clones are presented. The dotted horizontal and vertical line delineate the cutoff below 10 largest-volume clones in 1994 and 1997, respectively. |
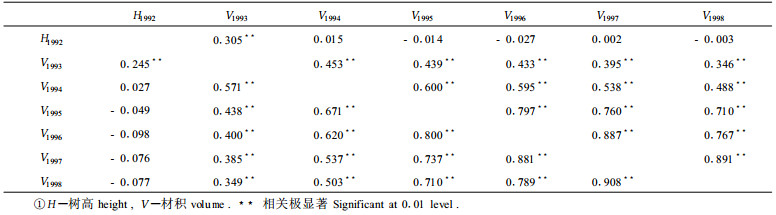
1992年无性系定植苗高与以后各年材积之间以及各年材积之间的普通相关(右上角)和等级相关(左下角)系数如表 5。表 5的数据表明,(1)定植苗高仅与第2 a的材积生长有较低的相关, 而与以后各年材积生长量之间没有相关。计算表明, 树高、直径的相关系数也有同样的表现。由于无性系苗系从湖南省运来, 上述事实可能是因为无性系与环境有较强的交互作用所造成, 即, 尽管会同种源优于京山种源, 但在湖南表现优良的无性系, 到湖北后表现并不一定好, 因此, 在湖北的试验是对无性系进行重新选择的过程。(2)1993年起至以后各年材积之间均存在极显著的相关, 但两种相关系数均随着年龄的增加而降低。“大群体”选择的效果决定于早期与伐期相关系数r的平方, 即决定系数r2(Wright, 1976)。上述结果预示着, 早期选择的效果是低的; 选择的时期越早, 效果越差。这一结果与前述表 2和表 3所得结论是一致的, 即, 早期表现的高增产率到25 a或30 a后采伐时将变得很小。
|
|
根据实际数据进行了早期选择少数无性系的模拟试验, 结果如图 2所示。若在1994年按材积选出10个最优无性系, 则到1997年时只有3个仍在前10名之内。这说明1994年保留的无性系数目太少, 以致把好的无性系淘汰了, 或者说, 它把许多并不优良的无性系选中了。有可能, 到25 a或30 a后采伐时, 它们之中没有1个可称作是“优良的”。从图中还可以看出, 如果1994年只选择4个无性系, 则到1997年时只有1个仍保留在前4名之内。
3 结论和讨论无性系林业是先进的林业生产技术。我国实行了近千年的杉木插条造林法被认为是世界上历史最久、规模最大的针叶树无性系林业(Bruce, 1992; Ritchie, 1994)。但是,我们现在却主要是采用种子繁殖。巳经知道, 采穗圃比种子园提早5~6 a提供苗木, 扦插苗比播种苗更高大粗壮, 成本更低, 而且,当用大群体繁殖时, 扦插苗造林与实生苗造林有相同的增产效果(李明鹤等, 1990a; Li Minghe et al., 1998;1999b)。本研究表明, 造林可以与无性系选择结合进行,由于去劣淘汰, 无性系林会比家系林有更高的生产力。因此, 作者认为, 杉木采穗圃应比种子园受到更多的重视, 可以由家系林业(family forestry)逐步走向无性系林业。
我国普遍认为传统的无性系育种法最适合于杉木,几个杉木5 a攻关计划均采用无性系育种方案。这种方法要经过至少一个轮伐期的无性系测验和区域化试验之后才可用于生产,需要很长的时间,不能为当前的林业生产服务。它不分好坏, 每个无性系都用很多的重复, 例如30株, 在同样的土地和经费条件下, 只能测定少数无性系, 例如200个, 很难选出真正优异的无性系。此外, 群体的遗传基础太窄, 难以适应山地复杂多变的环境和抵抗病虫害的侵袭。
本研究使用有增产效果的大群体, 再加上定期去劣, 可保证无性系群体在任何时候都可用于生产并有增产效果, 它把无性系选择与生产性造林相结合, 不需要特别的无性系测验和区域化试验阶段,因此,可以节约数十年的时间,直接为当前的造林提供大批苗木。此外,本方案采取了逐步增加重复次数的方法, 使得最优秀者得到最多的重复, 最差者得到最少的重复,这可以较少的经费和土地同时测定数千或数万无性系,因而可以选出十分优秀的无性系来。大量的无性系有利于林分在复杂多变的山地环境中保持产量稳定和抵抗病虫的侵袭。因此,作者认为,本方案比传统的无性系育种法更适合于杉木。
有些研究主张把仅仅经过了几年测验的少数无性系用于生产(马常耕, 1991; 周天相, 1990)。他们认为,一定采伐期的杉木无性系适宜的早期选择年龄为4~6 a (马常耕等, 2000;周天相等,2000)。本研究表明, 一次早期选择的增产效果很小,早期表现的高增产率是一种假象, 它会迅速下降, 到25 a或30 a后采伐时将变得很小。关于这一点,理论上早有明确的论证(李明鹤, 1990)。根据孔繁浩的研究(Kung, 1975)可以推断出,若在早期选出10%最好的无性系, 如早期与伐期的相关系数为0.3, 则到采伐时, 大约有82%的中选无性系已不在最好的10%中。此外,若在早期选出的是少数无性系,则由于遗传漂变,其结果将是不定的。本研究数据表明, 若在1994年选出10个最优无性系, 则到1997年时只有3个仍在前10名之内。这预示着, 到25 a或30 a后采伐时, 它们之中可能没有1个可称作是“优良的”。浙江开化林场1987年从3 a生的试验林中选出了4个优良无性系, 但到1998年时没有一个仍在前10名之内(周天相, 1990; 1998)。
胡伯智, 厉荣良, 冯建国. 1999. 杉木优良家系实生苗与扦插苗造林效果比较. 南京林业大学学报, (4): 74-75. DOI:10.3969/j.issn.1000-2006.1999.04.017 |
李明鹤. 1990. 遗传参数在林木早期选择中的应用. 湖北林业科技, (4): 1-4. |
李明鹤, 杨昌岩, 沈宝仙. 1990a. 大规模生产杉木改良苗木的方法. 林业科学, 26(4): 363-367. |
李明鹤, 李传涵, 杨通沂, 等. 1990b. 用杉木优树侧枝建立采穗圃技术. 林业科技通讯, (8): 27. |
李明鹤, 沈宝仙, 杨昌岩, 等. 1990c. 用杉木伐桩萌条建立采穗圃的研究. 林业科技通讯, (8): 14-15. |
李明鹤, 杉木无性系选择.见涂忠虞, 沈熙环编.中国林木遗传育种进展.北京: 科学技术文献出版社, 1993: 213-220
|
马常耕. 1991. 杉木无性系选育程序和苗木增殖技术. 林业科技通讯, (3): 5-7. |
马常耕, 周天相, 徐金良. 2000. 杉木无性系生长的遗传控制和早期选择初探. 林业科学, 专刊, (36): 62-69. |
周天相. 1990. 杉木无性系育种和良种繁殖新技术. 北京: 中国林业出版社, 38.
|
周天相. 1998. 杉木形质指标选择的效果. 林业科技通讯, (5): 6-7. |
周天相, 马常耕. 2000. 杉木无性系育种(开天系列). 林业科学, 36(3): 40-45. DOI:10.3321/j.issn:1001-7488.2000.03.007 |
朱之悌. 1986. 树木的无性繁殖与无性系育种. 林业科学, 22(3): 280-290. |
Bruce Zobel. 1992. Vegetative propagation in production forestry. Journal of Forestry, 90(4): 29-33. |
Kung F H. Efficiency of early or truncated selection. In: Proceedings of the Central States Tree Improvement Conference, 1975, 9: 35-41
|
Kleinschmit J, Schmidt J. 1978. Experiences with Picea abies cutting propagation in germany and problems connected with large scale application. Silvae Genetica, (26): 197-203. |
Li Minghe. Historical development of superior clones of Chinese Fir in China. In: Shen Xihuan. Forest tree improvement in the Asia-Pacific Region. Beijing: China Forestry Publishing House, 1995: 232-237
|
Li Minghe. The past and present of clonal forestry of Chinese Fir (Cunninghamia lanceolata (Lamb.) Hook.) in China. In: Jiro Kikkawa, Peter Dart, David Doley et al.ed. Proceedings of the 6th international workshop of BIO-REFOR. Brisbane, Australia, 1998: 84-87
|
Li Minghe, Ritchie G A. 1999a. Eight hundred years of clonal forestry in China: Ⅰ. Traditional afforestation with Chinese Fir (Cunninghamia lanceolata (Lamb.) Hook.). New Forests, 18(2): 131-142. DOI:10.1023/A:1006558900234 |
Li Minghe, Ritchie G A. 1999b. Eight hundred years of clonal forestry in China: Ⅱ. Mass production of rooted cuttings of Chinese Fir (Cunninghamia lanceolata (Lamb.) Hook.). New Forests, 18(2): 143-159. DOI:10.1023/A:1006553115255 |
Li Minghe, Shen Baoxian. Vegetative propagation and rejuvenation of Chinese Fir (Cunninghamia lanceolata (Lamb.) Hook.) in China. In: Jiro Kikkawa, Peter Dart, David Doley et al.ed., Proceedings of the 6th international workshop of BIO-REFOR. Brisbane, Australia, 1998: 122-123
|
Li Minghe, Shen Baoxian, Li Chuanhan. 1999c. Cloning strategy for Chinese Fir (Cunninghamia lanceolata (Lamb.) Hook.) suggested by early test results. Silvae Genetica, (48): 242-247. |
Lambeth C C. 1980. Juvenile-mature correlations in Pinaceae and implications for early selection. For. Sci, 26: 271-580. |
McKeand S E. 1988. Optimum age for family selection for growth in genetic tests of loblolly pine. For. Sci, 34: 400-411. |
Ritchie Gary A. Commercial application of adventitious rooting to forestry. In: Davis T D, Haissing B E ed. Biology of Adventitious Root Formation. New York: Plenum Press, 1994: 37-51
|