 2003, Vol. 39
2003, Vol. 39文章信息
- 方乐金, 施季森, 李力, 吴小龙, 史廷先.
- Fang Lejin, Shi Jisen, Li Li, Wu Xiaolong, Shi Tingxian.
- 枫香子代性状的遗传变异分析
- ANALYSIS OF GENETIC VARIATION OF PROGENY TRAITS IN LIQUIDAMBAR FORMOSANA
- 林业科学, 2003, 39(3): 148-152.
- Scientia Silvae Sinicae, 2003, 39(3): 148-152.
-
文章历史
- 收稿日期:2002-10-10
-
作者相关文章


2. 南京林业大学 南京 210037;
3. 安徽省祁门县林业局 祁门 245300
2. Nanjing Forestry University Nanjing 210037;
3. Forestry Bureau of Qimen County, Anhui Province Qimen 245300
自20世纪40年代以来,世界上许多国家注重短周期工业原料林的培育(方乐金,2000)。由于阔叶树的生产效益及产量不及针叶树,故许多国家在人工林发展初期,均把针叶树作为发展重点,中国亦不例外,如南方杉木、马尾松等一些针叶树种占总造林面积的80%以上(方乐金,2000)。大规模的针叶树替代了物种丰富的天然林群落,导致森林生态系统失衡,物种多样性降低,森林病虫害频频发生,人工更新难度加大等问题日益显露。同时,针叶树林产品相对过剩与阔叶树林产品不足,严重制约了林业自身的经济效益(方乐金,2000;施季森等,1996)。
中国枫香(Liquidambar formosana),春夏季节叶片呈嫩绿色,秋季叶片色泽五彩缤纷,适宜作园林景观树种;叶可提取芳香油,为“苏合香”的代用药品,作为皮肤杀菌药;树脂是塑料及人造革的原料;其锯屑、树梢是培养香菇、木耳的优良材料;木材轻软,质地坚实,纹理致密,易加工,旋切性能好,是建筑、家具和胶合板等的理想用材。枫香是我国南方林区中主要森林树种之一。树干圆满通直,枝叶繁茂,生长迅速。同时枫香适应性强,耐干旱瘠薄,易于天然更新,有荒山造林“先锋树种”之称。另外枫香抗污染能力强,尤其对SO2、氯化物有较强的抗性(《优良速生珍稀树种》编写组,1979;郑万均,1985;《中国香料植物栽培与加工》编写组,1986)。然而中国枫香主要性状遗传变异规律的研究几乎空白,开展中国枫香良种选育研究,不仅对丰富我国阔叶树造林树种,为工业用材提供原料,为园林绿化提供优质观赏树种,同时对改善林地生境、调整林业产业结构、改善生态环境等方面都具有重要的意义。
1 材料与方法 1.1 材料来源参试的枫香优树子代材料,来源于南京林业大学、安徽祁门县林业局、黄山学院合作开展的“中国枫香多目标育种及栽培技术研究”项目1996年选出的首批优树。1997年采集优树半同胞自由授粉种子,1998年育苗,1999年春季造林。参试子代82个,对照(CK)2个,计84个处理。按完全随机区组设计,4株小区,10次重复。2001年秋停止生长后,每木调查胸径、地径、全高、当年树高、冠幅、分枝数等性状及病虫危害状况。
1.2 统计分析对观测数据进行方差分析和协方差分析及多重比较测验等,以估算各生长性状的遗传参数,并分析它们的相关性(遗传相关、表型相关和环境相关)以及基因型与环境交互效应。采用SAS软件进行统计分析。
方差分析模型(莫惠栋,1983):

|
(1) |
式中,Yijk为第i家系在第j个重复的第k个观测值,Fi为家系效应,Bj为重复效应,(FB)ij为家系×重复的交互效应,εijk为机误。
遗传参数的估算(王明庥,1989):

|
(2) |

|
(3) |

|
(4) |
式中,hf2为家系遗传力,hi2为单株遗传力,δf2、δe2分别表示家系方差、机误,n为家系数, △G为遗传增益,S为选择差,h2为广义遗传力,i为选择强度,δp为性状表型标准差。
相关分析(王明庥,1989):
用小区平均值对每一性状(x, y)进行协方差分析:

|
(5) |

|
(6) |
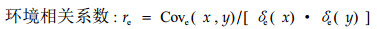
|
(7) |
式中,Cove(x, y)、Covf(x, y)、Covp(x, y)分别为x性状与y性状的环境协方差、家系基因型协方差和表型标准差协方差;δe(x)与δe(y)、δf(x)与δf(y)、δp(x)与δp(y)分别为x性状与y性状的环境方差、家系方差和表型标准差。
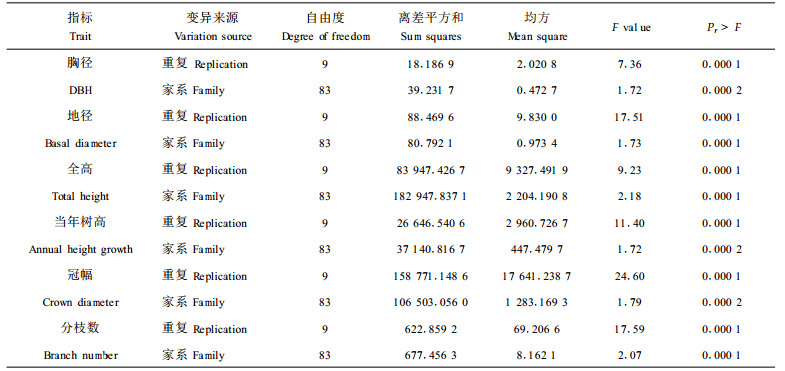
2 结果与分析 2.1 枫香子代生长性状的方差分析各家系主要生长性状的方差分析结果表明,3 a生枫香子代全高、当年树高、地径、胸径、冠幅、分枝数等性状差异均达极显著水平(见表 1)。
|
|
枫香子代测定林各生长性状的遗传力估算结果(表 2)表明,枫香在幼林期,各性状受中度、中低度遗传控制。其中树高性状遗传力估值最高,家系达到0.54,单株达0.43,分枝数次之,地径、胸径、当年树高、冠幅等性状遗传力相近,在0.42~0.44之间。表型变异系数和遗传变异系数为树高最小,分别为16.80%和16.38%,表明树高性状遗传较稳定。
|
|

遗传相关系数是研究数量性状相关变异的一个重要参数,在间接选择和指数选择等方面广泛应用。本研究结果(表 3)表明,全高与当年树高、胸径与地径的遗传相关系数较大;分枝数与其他各性状间的遗传相关系数较小,表明分枝数性状具有较强的独立遗传能力;其余各性状间遗传相关程度中等。
|
|
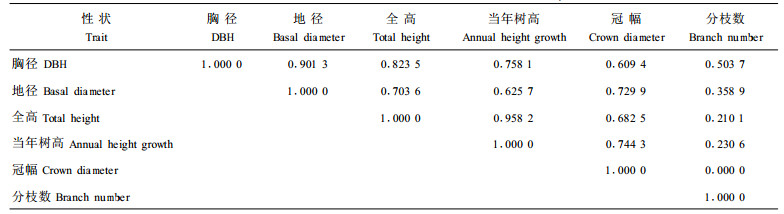
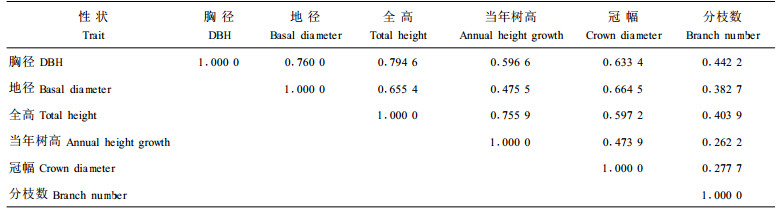
表 4表明,枫香子代测定林幼林期,全高与当年树高性状间表型相关系数较大,生长量存在显著的正相关。全高同时与胸径表型相关系数亦呈显著正相关,这种表型相关随树龄增长、林分开始分化后,将会有所减弱。分枝数其他性状间表型相关系数相对较小,尤其是与当年树高的相关系数最低,说明枫香分枝数的多少与其它性状相关较弱。其余各性状间表型相关系数亦较大,但未达显著水平。
|
|
全高与胸径、当年树高间的环境相关系数较大(表 5),说明这些性状对环境变化敏感,立地条件好则生长量大,反之则小。而分枝数与冠幅等的环境相关系数较小,表明它们对环境条件较为迟钝。
|
|

通常用材林树种以其最终获取的木材产量作为首要指标,结合干形、抗性、适应性等性状选择优良家系。因本研究的子代测定林尚处幼林阶段,材积遗传性状未能充分表达, 故以构成材积的树高和胸径为指标,且遗传增益大于5%为选择标准,对参试家系进行了初步选择(表 6),有118#、58#、113#、70#、119#、123#6个家系树高、胸径的遗传增益幅度都较大,而101#、29#、60#3个家系,胸径分别达7.86%、7.74%、5.54%,而树高分别仅为3.78%、2.67%、2.35%,胸径遗传增益幅度高于树高107.9%~188.8%。
|
|

首批参试的84个枫香子代,3 a生时全高、胸径、地径、当年树高、冠幅、分枝数各性状差异均达显著以上水平。家系遗传力和单株遗传力估值以全高、分枝数2个性状较高,其它性状较低。枫香子代家系间存在丰富的遗传变异,具有较大的遗传改良潜力。相关性分析表明各性状间遗传相关、表型相关、环境相关均存在一定的相关关系,为开展枫香选择和改良提供间接选择的理论依据。以树高和胸径为指标,且遗传增益大于5%为选择标准初选的9个优良家系,有6个家系全高、胸径的遗传增益幅度较高,3个家系全高、胸径的差异较大。但受测定林年幼的影响,各性状的基因表达未能充分表现,各项遗传参数未趋于稳定,有待测定林发育到一定阶段才能作出较可靠的结论。
方乐金. 2000. 枫香选优技术的探讨. 浙江林业科技, 20(5): 47-49. DOI:10.3969/j.issn.1001-3776.2000.05.013 |
莫惠栋. 1983. 农业试验统计. 上海: 上海科学技术出版社, 259-278.
|
施季森, 朱胜利, 马如兴, 等. 1997. 中国枫香多目标育种的研究Ⅰ中国枫香优树选择. 林业科技通讯, (10): 15-16. |
王明庥主编.林木育种学概论.北京: 中国林业出版社, 1989: 191-207
|
《优良速生珍稀树种》编写组. 1979. 优良速生珍稀树种. 第一版. 南昌: 江西人民出版社.
|
郑万均主编.中国树木志.第二卷.北京: 中国林业出版社, 1985
|
《中国香料植物栽培与加工》编写组. 1986. 中国香料植物栽培与加工. 北京: 中国林业出版社.
|
SAS Institute Inc. 1990. SAS Fundamentals:A Programming Approach. SAS Campus Drive, Cary, NC27513. |