 2003, Vol. 39
2003, Vol. 39文章信息
- 张建国, 盛炜彤, 罗红艳, 熊有强.
- Zhang Jianguo, Sheng Weitong, Luo Hongyan, Xiong Youqiang.
- 杉木营养平衡与苗木干物质的分配关系
- STUDIES ON RELATIONSHIP BETWEEN NUTRITION BALANCE AND ALLOCATION OF PHOTOSYNTHETIC PRODUCTS IN SEEDLINGS OF CHINESE FIR
- 林业科学, 2003, 39(3): 37-44.
- Scientia Silvae Sinicae, 2003, 39(3): 37-44.
-
文章历史
- 收稿日期:2002-11-13
-
作者相关文章


2. 中国林业科学研究院亚热带林业实验中心 分宜 336600
2. Experimental Centre of Subtropical Forestry, CAF Fenyi 336600
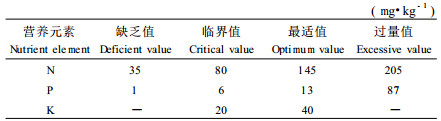
树木营养亏缺是由于土壤的营养元素不足和养分的不平衡造成的。关于营养诊断,目前主要采用临界值法和综合诊断法(DRIS法)。临界法的缺陷在于仅以单一营养元素的亏缺状态为诊断标准,难以判断营养平衡的状态;而DRIS法(Beaufils, 1973)的优点是能给出平衡态时的各营养元素之间的比例,缺点是确定的平衡态往往是低水平的,而非真正的平衡。目前关于营养诊断方法的合理性仍然缺乏理论上的严密论证。关于杉木的营养与施肥效应已有许多研究(李贻铨等,1992;范少辉等,1996;张建国等,1996),但是对营养诊断和营养平衡仍缺乏深入的探讨。本文系统研究了杉木根冠功能平衡与营养平衡的关系,从苗木对营养胁迫的适应机理及营养胁迫与杉木苗木光合产物分配模式的关系角度出发,建立了苗木营养平衡理论,提出了一种全新的群体营养诊断方法,给出了杉木的营养诊断标准,并通过大量杉木人工林施肥试验证明了这一方法的正确性。
1 材料和方法 1.1 材料参加试验的苗木类型为杉木当年生播种移栽芽苗、1 a生裸根苗和1 a生扦插苗。供试土壤为黄红壤。1 a生扦插苗供试土壤取自树木园0~40 cm土层(A层),捣碎后加20%的珍珠岩混匀而成培育基质,土壤pH为5.5,有机质6.4 g·kg-1,全N为1.24 g·kg-1,有效N为119 mg·kg-1,速效K为65 mg·kg-1,有效P为1.77 mg·kg-1。移栽芽苗和1 a生裸根苗供试土壤取自树木园40~100 cm土层(B层),经捣碎混匀而成培育基质,土壤pH为5.02,有机质6.12 g·kg-1,全N为0.54 g·kg-1,有效N为40.46 mg·kg-1,速效K为47.88 mg·kg-1,有效P为1.81 mg·kg-1。由于0~40 cm土层和40~100 cm土层土壤在有效P含量方面差异不显著,所以培育基质的差异主要在N素营养的差异上。但从整体看,0~40 cm土壤培育基质处于P营养亏缺状态,N、K相对比较充足。40~100 cm土壤培育基质处于N、P营养亏缺状态,而K素较为充足。因此,0~40 cm土层土壤可作为研究P营养亏缺的培育基质,40~100 cm土层土壤可作为研究N、P营养亏缺的培育基质。
1.2 试验设置在N、P素营养亏缺的培育基质上,共设计了N、P、NP 3种营养试验。
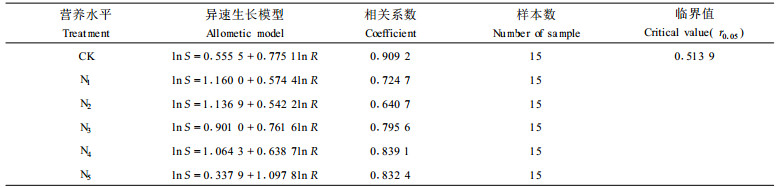
N营养试验共设计了6个供N水平,选用肥料为尿素(含有效N 46%),6个处理分别为CK(对照)、N1(1 g·盆-1)、N2(2g·盆-1)、N3(3g·盆-1)、N4(4g·盆-1)、N5(5g·盆-1),参试苗木为当年生移栽芽苗(1 a后测定时已为1 a生裸根苗)和1 a生播种裸根移栽苗(1 a后测定时已为2 a生裸根苗)。试验容器采用黑色塑料桶,规格为直径31 cm,高22 cm,每盆装填土10 kg。芽苗每盆定植5株,1 a生苗定植4株。
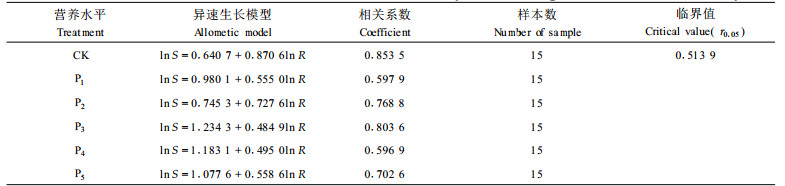
P素营养试验设计与N营养试验设计相同, 肥料选用一级钙镁磷肥。6个处理分别为CK(对照)、P1(2g·盆-1)、P2(3g·盆-1)、P3(4g·盆-1)、P4(5g·盆-1)、P5(6g·盆-1)。
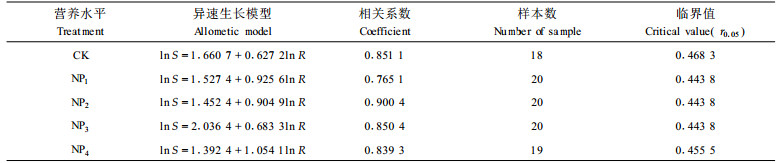
NP营养试验共设计了5个供NP水平,选用肥料为(NH4)2HPO4,5个处理分别为CK(对照)、NP1(0.02%)、NP2(0.04%)、NP3(0.08%)、NP4(0.12%),百分数表示单位鲜土的施磷量。参加试验的苗木为1 a生扦插苗(测定时已为2 a生),试验容器采用泥盆,规格为直径28 cm,高19 cm。每盆装土7.5 kg,每盆定植4株。
1.3 试验期苗木的管理苗木定植后,浇足底水,以后定时定量浇水。定植后的苗木在温室缓苗1个月,然后将苗木移到室外防雨棚下约10 d左右,然后开始施肥处理,并进行细致管理。施肥处理方法是先定量称取肥料,用水溶解稀释后浇入,肥料分2次施入,间隔时间为15 d。
1.4 指标测定苗木在定植5 d后开始测定记录,1 a生苗木试验测定苗高和地径,芽苗试验只测定苗高。在施肥处理的前1 d,全部测定一次苗木地径、苗高,所有处理在施肥后每隔10 d测定一次。试验结束后将苗木全部挖出,洗净,分株测定根、茎、叶生物量。
2 结果分析 2.1 营养平衡理论的建立张建国等(2003)研究表明,不同营养胁迫条件下,杉木苗木光合产物累积分配的模式是不同的。在N、P亏缺状态下,增加N素的供应,导致2 a生苗光合产物的累积由根、茎向叶部转移;对1 a生苗,施P使得光合产物的累积重点由根系向茎部和叶部转移;与1 a生苗木施磷不同,2 a生苗木施磷使得光合产物累积由茎向根部转移。在N素营养充足、P素亏缺状态下,施N P肥对苗木干物质分配规律的影响是:随着P营养亏缺的解除,光合产物累积由根向茎、叶转移,其中向茎转移累积量为0.5%~3.13%,向叶部转移为7.21%~11.76%,可见光合产物累积主要向叶部转移。
光合产物的不同分配模式其实质反映了苗木对营养胁迫的适应性反应。关于这一点,Davidson(1968), Hunt(1975), Thornley(1972a;1972b)等人从不同角度曾论述过一种机制,即当土壤因子(营养或水)受限制时,大部分水或营养物质被用于根系生长,以增加根冠比(R/S)。后来在此基础上,Brouwer(1983)提出根冠功能平衡的思想,即植物根、冠功能是互补的,冠层的功能是进行光合作用,合成碳水化合物,根系的功能是吸收营养和水分,当其一功能下降时,它们便相互竞争各自所需物质,植物生长所需物质优先供应近源组织生长。植物体内有自动调节功能,最终使根、冠向减少伤害、适应环境胁迫的方向发展,从而达到一种新的平衡状态。
根据营养胁迫与杉木苗木光合产物分配的关系,提出如下营养平衡理论:
2.1.1 光合产物分配模式(1) 在营养亏缺条件下,苗木光合产物累积由冠部向根部转移,累积速率根部大于冠部,从而使得R/S提高。R/S的提高有利于苗木吸收更多的营养和水分,以维持苗木在营养亏缺条件下的生存和生长,是苗木对营养亏缺逆境的适应性反应,但苗木仍处于非平衡状态,整体生物量比较低。当外加营养后,如果苗木的光合产物的累积由根向冠部转移,R/S下降,表明苗木的营养亏缺得到了一定改善或解除;反之,如果苗木的光合产物由冠向根部转移,R/S上升,表明苗木的营养亏缺不仅没有得到改善,反而进一步加剧了苗木营养失衡。(2)在营养平衡条件下,苗木光合产物在冠部和根部平衡累积,其累积速率相等,R/S维持在某一较低的而稳定的水平上,表明苗木处于与营养环境最为协调的平衡状态,干物质产量也达到最大。要指出的是平衡态的营养往往具有一定的范围,非某一固定值。(3)在一定营养过量条件下,苗木光合产物累积仍由根部向冠部转移,累积速率冠部大于根部,从而使得根冠比R/S维持在某一较低的范围内。当营养过量超过一定限度后,由于营养平衡严重失调,过量的营养成为毒害因子,从而导致苗木干物质产量开始大幅度下降,并且随着营养失调的进一步加剧,苗木开始死亡。一般,在自然状态下营养过量的情况是很难出现的,但人为导致的营养过剩时有发生。如农作物施肥过量的情况下引起地上部分徒长,易倒伏,造成减产就是一个很好的例证。

|
图 1 不同营养条件下苗木光合产物的累积方向 Fig. 1 Movements of photosynthetic products in seedlings under different nutrient conditions |

|
图 2 不同营养条件下干物质产量与根冠比R/S的变化规律 Fig. 2 Change patterns of dry matter and root/shoot ratios in seedlings under different nutrient conditions |
根冠干物质分配的关系可应用异速生长模型y=axk来描述。为了明确表示根冠相互关系,采用如下表达式:ln S = a + k ln R,式中,R表示根系干物质重,S表示冠部干物质重,k表示根冠生长速率的关系参数。
2.1.3 营养平衡诊断法则根据营养胁迫与苗木干物质分配的关系及异速生长模型参数k值对营养胁迫反应的敏感性变化规律,提出如下新的营养诊断法,称之为群体营养诊断法,其判别法则为:(1)在某一营养条件下,当苗木参数k<1时,表明根系相对生长速率高于冠层相对生长速率,苗木处于营养亏缺状态;(2)当k>1时,表明苗木冠层相对生长速率高于根系,苗木土壤某一营养物质供应已开始过剩。一般在自然状态下,k>1是不会出现的。(3)当k=1时,表明根系和冠层相对生长速率相等,苗木处于营养平衡状态。
2.2 营养胁迫与苗木干物质的分配的关系表 1~3是不同营养胁迫条件下苗木干物质分配模式的变化规律拟合结果。从表 1~3可以明显看出,异速生长模型中参数a、k对不同营养水平(N、P、NP)的反应是非常敏感的,而且具有明显的变化规律。以不同N营养水平对杉木2a生苗木生长影响试验为例,对照参数k值为0.775,a值为0.555;N1~N4 4个处理参数k值下降,为0.542~0.761,a值上升,为0.901~1.166;N5处理k值最大,为1.09,a值下降,为0.3379。
|
|
|
|
|
|
在不同P营养水平对杉木2a生苗木生长影响的试验中,对照参数a为0.6407,随着P的施入a值增加,为0.7453~1.2345,k值的变化与a值正好相反,对照k最大,为0.8706,施P后k值下降,为0.4849~0.7276。
在不同NP营养水平对杉木2a生苗木生长影响的试验中,a、k的变化均不同于N或P处理试验,由于供试基质N素充足,不同NP施肥处理参数a值为1.3942~1.5247,均低于对照(1.660),但k值(0.6833~1.0541)均高于对照(0.6272)。
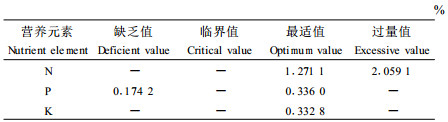
2.3 营养平衡诊断对表 1~3的方程进行绘图(见图 3~5)。根据以上判别法则,从图 3~5可以明显看出,N肥试验的对照(CK)和N1~N4处理其方程均偏离平衡线k=1,所以苗木仍处于营养亏缺状态,N5处理k值为1.09,接近平衡线,表明苗木处于营养平衡状态;施P的处理试验,由于供试基质N素亏缺,苗木同样处于营养亏缺状态,施P后反而导致苗木k值下降,营养平衡进一步失调,由此可以看出平衡施肥的重要性;NP试验由于供试基质N素充足,NP1、NP2、NP4 3个处理营养接近平衡线,表明苗木处于营养平衡状态,而CK和NP3处理仍然处于亏缺状态。根据以上分析,依据营养平衡判别法则,进一步可提出杉木苗木营养诊断标准,见表 4~5所示。

|
图 3 不同N营养条件下2 a生苗木异速生长模型 Fig. 3 Changes of Allometic models of 2-year-old seedlings under the different N nutrient status N1与N22线近乎重叠 Two lines of N1 and N2 nearly overlap. |

|
图 4 不同P营养条件下2 a生苗木异速生长模型 Fig. 4 Changes of Allometic models of 2-year-old seedlings under the different P nutrient status |

|
图 5 不同NP营养条件下2 a生苗木异速生长模型 Fig. 5 Changes of Allometic models of 2-year-old seedlings under the different NP nutrient status |
|
|
|
|
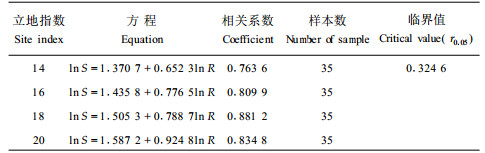
为验证上述营养判别法在野外应用的可行性,在中国林业科学研究院亚热带林业实验中心年珠林场进一步做了野外造林试验,造林苗木为1 a生裸根苗(试验测定时已为2.5 a生),造林试验地设置在同一坡向上,分4个立地指数(14、16、18、20)小区,面积0.27 hm2。1997年进行苗木干物质分配模型调查,调查时苗木年龄2.5 a。异速生长模型方程拟合结果见表 6和图 6。从表 6和图 6可以明显看出,18指数(含18指数,下同)以下苗木干物质分配模型参数k为0.6523~0.7887,均小于1,表明18指数以下立地苗木处于营养亏缺状态,并且随立地指数的下降,亏缺越严重,而20指数立地k值为0.9248,接近平衡线,苗木处于营养平衡状态。由此可以确定,杉木幼林施肥的有效立地指数区间为10~18。并且,施肥效应随立地指数的上升而下降。关于这一点将从施肥试验的角度进一步分析和证明。
|
|

|
图 6 不同立地指数幼林2.5 a生苗木根冠异速生长模型的变化 Fig. 6 Changes of Allometic models of 2.5-year-old seedlings with different site indexes |
此外,通过营养胁迫试验,发现参数k与苗木根冠比(R/S)之间的关系存在一定规律,可以用k=a+bln(R/S)来表示,结果如图 7~10所示。图 7~10表明,盆栽苗木的R/S值在0.6~1.0之间,以k=1进行判别,营养平衡时2a生苗木的根冠比为0.6;野外试验地苗木R/S值在0.2~0.4之间,k=1时2.5a生苗木根冠比为0.2。根据R/S与k值的关系,同样可简便地对苗木的营养平衡状态进行判别。

|
图 7 不同N营养条件下k与R/S的关系 Fig. 7 Relationship between k and R/S under the different N nutrient conditions |

|
图 8 不同P营养条件下k与R/S的关系 Fig. 8 Relationship between k and R/S under the different P nutrient conditions |

|
图 9 不同NP营养条件下k与R/S的关系 Fig. 9 Relationship between k and R/S under the different NP nutrient conditions |

|
图 10 不同立地指数幼林2.5 a生苗木k与R/S的关系 Fig. 10 Relationship between k and R/S of 2.5-year-old seedlings with different site indexes |
施肥的目的主要是补充土壤养分不足,提高林木的生长量。表 7的结果说明,杉木幼林施肥肥效随立地指数的变化而变化。在10~12指数立地,土壤肥效反应最大,其次为14~16指数,18指数已无明显肥效。这一规律与不同立地指数异速生长模型中k值反应的规律是完全一致的,这进一步证明了本文提出的k值作为营养平衡判别方法的正确性。对于杉木幼林施肥肥效反应的规律可以应用土壤营养诊断标准(表 5)来解释。从表 7可以看出,河南信阳试验点,立地指数为10~12,有效N、P,速效K含量为64.2 mg·kg-1、3.84 mg·kg-1、18.21 mg·kg-1,均低于临界值,表明林地土壤处于营养亏缺状态,因此施肥以N100P100K100最大,肥效反应顺序为N>P>K;江西分宜试验点立地指数为14,有效N、P,速效K含量为258.3 mg·kg-1、1.87 mg·kg-1、85.07 mg·kg-1,按营养诊断标准,林地土壤N素和K素充足,P素亏缺,因此施肥以P素肥效最大,施N反而有负作用,如N100 P100K100处理树高反而低于P100K100;广东始兴立地指数为16,有效N、P,速效K含量分别为114.4 mg·kg-1、2.78 mg·kg-1、73.99 mg·kg-1,根据营养诊断标准,林地土壤N素亏缺、P素极度亏缺、K素充足,因此P素肥效最大,其次为N素,K素无效;福建顺昌试验点立地指数为18,有效N、P,速效K含量分别为151.07 mg·kg-1、3.95 mg·kg-1、53.34 mg·kg-1,根据营养诊断标准,林地土壤N素和K素充足,P素亏缺,因此P素有一定肥效,N素无效或有负效应,K素无效。
|
|
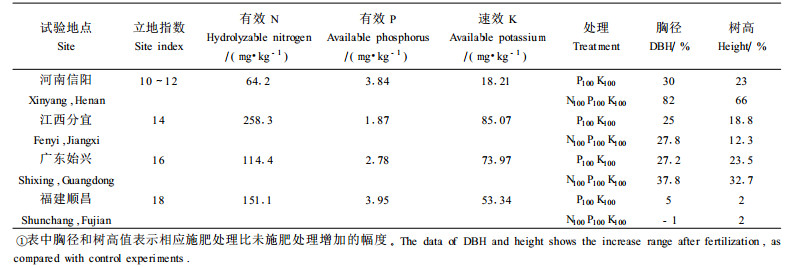
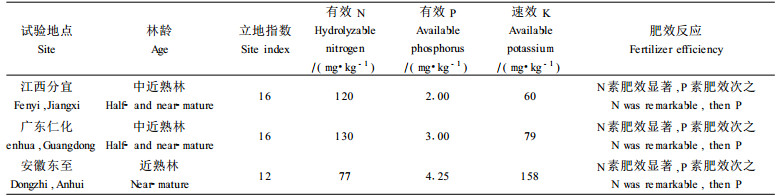
表 8表明,中近熟林施肥,N素肥效最大,其次是P素,K素无肥效。供试的所有的结果表明,以NP肥效最大。中近熟林施肥的肥效反应规律表明,杉木在进入中龄林后,由于树木营养需求量增大,加之立地养分的消耗,林木开始发生营养亏缺,尤其是对N素的需求量增大而导致的林地土壤N素的亏缺增大,这一点与幼林完全不同,反应了杉木中近熟林的营养需求特性。表 5的土壤营养诊断标准与表 8的施肥试验结果比较,很明显可以看出,立地土壤N、P处于亏缺状态,因此施N、P有明显效果,但相比较而言,P素肥效低于N素。关于P素肥效比N低的原因,笔者认为这主要决定于林木对P的绝对需求量,对进入中龄以后的杉木来说,林木体内P的绝对含量已达到临界水平以上,再加上从老叶转移到新叶的P,相对幼林而言,中近熟林林木对P的需求量相对下降,因此施P的肥效也低于幼林。
|
|
关于中近熟林林下土壤营养状况,在福建杉木中心产区对19块林分立地土壤肥力状况测定(0~40 cm)的结果如图 11所示。从图 11可以明显看出,中近熟林林分土壤肥力N、P处于营养亏缺状态,而K素比较充足。在10~16指数立地范围内,有效N含量在40~110 mg·kg-1,并且N素含量随立地指数的下降而下降;有效P含量在0.1~2.0 mg·kg-1范围内,其变化与立地指数无明显关系;速效K含量为40~140 mg·kg-1,比较高,随立地指数的下降而下降;根据表 5土壤营养诊断指标标准,N、P处于低水平或缺乏状态,施N素和P素肥效必然显著,而施K素无肥效。

|
图 11 16~18 a生中近熟林有效N、有效P、速效K含量与立地指数的关系 Fig. 11 Relationship between the content of hydrolyzable nitrogen, available phosphorus and available potassium and site indexes in the 16~18 years old near-mature forest |
苗木根冠干物质分配的关系可应用异速生长模型y=axk来描述。由于不同营养胁迫可导致苗木光合产物的分配模式发生明显改变,因此异速生长模型可应用于苗木的营养研究,来描述苗木的营养状态。试验表明,异速生长模型参数k值对营养胁迫反应非常敏感,k值实际上就反映了苗木的营养状态。依据k值进行营养状态的判别方法是:在某一营养条件下,当苗木参数k<1时,表明根系相对生长速率高于冠层相对生长速率,苗木处于营养亏缺状态;当k>1时,表明苗木冠层相对生长速率高于根系,苗木土壤某一营养物质供应已开始过剩,一般在自然状态下,k>1是不会出现的;当k=1时,表明根系和冠层相对生长速率相等,苗木处于营养平衡状态。依据k值的变化规律和Brouwer(1983)对根冠功能平衡的解释,建立了苗木营养平衡理论,提出了新的营养诊断方法。
根据新的营养诊断方法,制定了杉木叶片和土壤营养诊断标准,并通过大量的施肥试验结果证明了诊断标准的正确性,同时也证明了新的诊断方法的可行性。
众所周知,树木的生物产量取决于总干物质生产及其在各个器官之间的分配,如何科学而合理地调节根冠比,使光合产物能更多地分配到目标产量部分,是一个很有价值的研究课题。本文系统地研究了酸性黄红壤上不同N、P、NP营养对杉木苗木根冠比的干物质分配模型的影响,揭示了杉木苗木营养需求特性及其对营养亏缺的适应机制,为苗木的培育、林木施肥、高效营养基因型树种的选择提供了理论依据。
范少辉, 俞新妥. 1996. 杉木人工林栽培营养的研究. 林业科学研究(杉木人工林栽培营养的研究专刊), 9(专): 1-9. |
李贻铨, 徐清彦. 1992. 杉木幼林前5年施肥效应研究. 土壤通报, 22(1): 28-32. |
张建国, 李贻铨, 纪建书, 等. 1996. 杉木幼林施肥的时效性研究. 林业科学研究(林木施肥与营养专刊), 9(专): 27-33. |
张建国, 盛炜彤, 罗红艳, 等. 2003. N、P、NP营养对杉木苗木生长和光合产物分配的影响. 林业科学, 39(2): 21-27. DOI:10.3321/j.issn:1001-7488.2003.02.004 |
Beaufils E R. 1973. Diagnosis and recommendation integrated system(DRIS). Soil Bulletin, 1. |
Brouwer R. 1983. Functional equilibrium: sense or nonsense?. Neth J Agric Sci, 31: 335-348. |
Davidson R L. 1968. Effect of root/leaf temperature differentials on root/shoot ratios in some pasture grasses and clover. Ann Bot, 33: 561-569. |
Hunt R. 1975. Root/shoot equilibrium in cranberry(Vaccinium macrocarpon Ait). Ann Bot, 39: 807-810. DOI:10.1093/oxfordjournals.aob.a084996 |
Thornley J H M. 1972a. A balance quantitative model for root/shoot ratios in vegetative plants. Ann Bot, 36: 431-441. DOI:10.1093/oxfordjournals.aob.a084602 |
Thornley J H M. 1972b. A model to describe the partitioning of photosynthesis during vegetative plant growth. Ann Bot, 36: 419-430. DOI:10.1093/oxfordjournals.aob.a084601 |
Thornley J H M. 1976. Mathematical models in plant physiology. London: Academic Press.
|