 2003, Vol. 39
2003, Vol. 39文章信息
- 侯祝强, 姜笑梅, 骆秀琴, 江泽慧, 费本华.
- Hou Zhuqiang, Jiang Xiaomei, Luo Xiuqin, Jiang Zehui, Fei Benhua.
- 针叶树木材细胞力学及纵向弹性模量的计算——试件纵向弹性模量的预测
- MECHANICS OF WOOD CELL AND ESTIMATION OF LONGITUDINAL ELASTIC MODULUS OF SOFTWOOD———ESTIMATION OF LONGITUDINAL ELASTIC MODULUS OF SPECIMEN
- 林业科学, 2003, 39(2): 123-129.
- Scientia Silvae Sinicae, 2003, 39(2): 123-129.
-
文章历史
- 收稿日期:2000-10-24
-
作者相关文章


本文是“针叶树木材细胞力学及纵向弹性模量的计算———纵向弹性模量的理论模型”(江泽慧等, 2002) (以下简称理论模型)工作的继续, 将应用前一部分工作中已获得的有关模型、计算公式及方法, 根据针叶树木材细胞的解剖结构参数, 计算试件的纵向弹性模量并与测定结果进行比较, 同时预测试件宏观弹性模量随细胞解剖结构的变化, 并分析讨论两者间存在着的定量关系和规律。
1 纵向弹性模量计算参数的确定 1.1 管胞解剖结构参数欲计算管胞整体的纵向弹性模量, 首先必须确定计算中所要使用的管胞解剖结构参数:管胞的长度、径向和弦向直径、径向和弦向壁厚度。试件包含有数目巨大的管胞, 而且这些管胞的大小各异, 无法也不可能对这所有的管胞一一进行测定。为此, 本文使用计算机随机模拟的方法, 确定试件中每一个管胞的上述5个解剖结构参数。姜笑梅(1995)和侯祝强等(2001)已经证明, 马尾松管胞的长度遵从随机正态分布; 同时, 侯祝强等(2001)还利用均匀分布随机数发生器对马尾松管胞长度进行计算机随机模拟, 模拟所得结果具有的精度完全可以满足科研与生产的要求。此外, 其它的一些研究工作已经证明, 某一些针叶树木材管胞的径向和弦向直径也服从随机正态分布(Vysotskaya et al., 1989)。因此, 本文使用正态分布函数来模拟试件管胞长度、径向和弦向直径、径向和弦向壁厚度。
在正态分布的前提下, 关于管胞解剖结构参数的计算机随机抽样可分为3个基本步骤(高惠璇, 1995; 侯祝强等, 2001): (1)运用样本参数的平均值和方差估算总体的平均值和方差, 构成管胞解剖结构参数的正态分布密度函数; (2)利用均匀分布随机数发生器实现标准正态分布的随机数的计算机抽样; (3)由解剖结构参数的样本平均值和标准差, 以及计算机抽样所得的标准正态分布随机数, 求出相应解剖结构参数的模拟值。
姜笑梅等(1994;1995)曾对人工林杉木(Cunninghamia lanceolata)、马尾松(Pinus massoniana)幼龄材和成熟材的解剖性质进行了研究, 而骆秀琴等(1995a;1995b)又对来自同一地点的人工林杉木、马尾松幼龄材和成熟材的力学性质进行了测定和研究。为便于将计算结果与测定结果进行比较, 本文选取这2种针叶树木材作为研究对象。采用姜笑梅等(1994;1995)关于人工林杉木、马尾松幼龄材和成熟材解剖结构参数的有关数据, 计算和预测这4种试件的纵向弹性模量; 再利用骆秀琴等(1995a;1995b)关于相应气干试件在常温条件下纵向弹性模量的测定值, 验证计算结果的准确性及所提出的针叶树木材细胞模型、试件纵向弹性模量计算方法的正确性。
表 1中列出的关于管胞的数据, 均引自前述的姜笑梅等两篇研究论文, 包括人工林杉木、马尾松幼龄材和成熟材的早晚材管胞长度的平均值与标准差、径向和弦向直径的平均值与标准差、径向和弦向壁厚度的平均值与标准差。此外, 利用与姜笑梅等研究工作所用试材同一地点的试材制作试样, 另外测定了人工林杉木、马尾松管胞早材与晚材的搭接率, 其平均值分别为25.3%和26.4%。
|
|
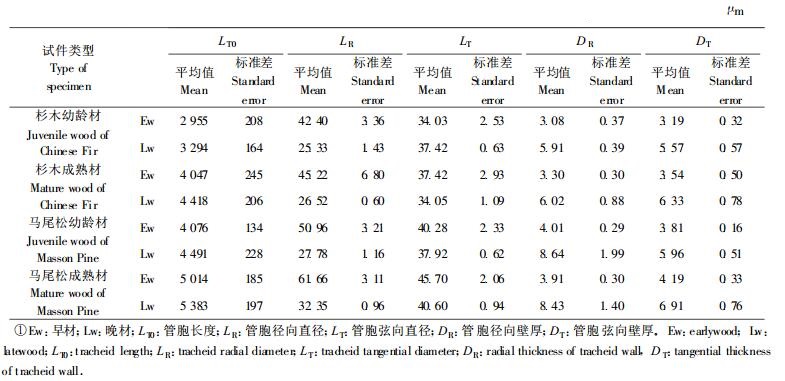
如本研究的理论模型所述, 由于无法对射线细胞的解剖结构参数进行计算机抽样模拟, 本文使用射线细胞有关的解剖结构参数平均值计算其纵向弹性模量。针叶树木材的射线管胞长度是管胞1/30(北京林学院, 1983), 平均为0.1~0.2 mm, 而其长度是其宽度的5~10倍。作为一个平均的估算, 本文对于射线细胞不作早晚材的区别, 取射线细胞长度为相应早材管胞长度的1/30, 取其直径为早晚材管胞径向和弦向直径的平均值; 另外, 射线主要由薄壁细胞组成, 取射线细胞胞壁为相应早材管胞壁厚度的1/10。表 4列出射线细胞纵向弹性模量的计算结果。
1.3 MP层和3S层的弹性模量沿用Mark等(1967; 1970)、Gillis(1970)关于细胞壁的假定:管胞胞壁的厚度和弹性模量与树种无关, 只在早晚材管胞间存在差异, 而MP层和3S层沿纤维方向和垂直纤维方向的弹性模量不相同, 其具有平面各向同性(plane isotropy)的特性。同时, MP层和3S层的厚度及其弹性模量采用Mark和Gillis (1970)研究论文的数据(表 2)。表中所列MP层的“E∥”和“E⊥”与本研究理论模型有关计算式中的“E1M”和“E2M”相对应, 而3S层的“E∥”和“E⊥”则与本研究理论模型有关计算式中的“E1S”和“E2S”相对应。
|
|

骆秀琴等(1995a;1995b)运用“四点加载”法及计算公式, 测定了人工林杉木、马尾松幼龄材和成熟材的抗弯弹性模量MOE。Bodig等(1982)指出, 可由“四点加载”测量抗弯弹性模量的方法得到纵向弹性模量EL, 因为试件的MOE与EL间存在如下的一个关系:

|
(1) |
式中: b为试件两个支撑点之间的跨度, h为试件沿负载方向的厚度。骆秀琴等关于试件MOE的测定按国家标准进行(GB1927-43-91, 1991), 可知b =300.00 mm及h =20.00 mm。表 4中所列出的试件测定值, 即根据骆秀琴等关于试件MOE的测定结果和试件的b、h值, 运用(1)式计算得出。要指出的是, 以往已有过关于利用弯曲实验的方法, 测定木材3个纹理方向弹性模量的报道(Watanabe, 1998)。
为了便于比较, 4种试件的早材、晚材、早晚材管胞纵向弹性模量的计算值, 也在表 4中同时一并列出。同时, 根据测定值的有效数位, 所有计算结果均取4位有效数字; 为使表中数字书写简洁, 各弹性模量的单位为GPa(1GPa=1×109 Pa)。
2.2 试件纵向和总的管胞数目如果试件纵向长L、横截面面积为A, 则试件纵向第i列所排列的管胞数目N1i为
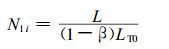
|
(2) |
而在R-T平面(试件的横截面)上, 管胞排列的数目NRT为

|
(3) |
由于各个管胞的长度不相同, 因此试件中纵向各列的管胞数目N1i不为常数, 根据计算机抽样模拟的结果, 其差异在1~2之间。这样的差异只占纵向各列管胞数目的1/40~1/50, 利用串联管胞纵向弹性模量公式计算可知(本研究理论模型中的(20)式), 其对于纵向弹性模量计算结果不产生有意义的影响。鉴于一个试件纵向弹性模量的计算过程中, 抽样模拟及其计算涉及到的管胞数目巨大, 在9.2×106~4.1×107的范围内, 完成一次计算耗时不短。为简化计算过程缩短计算时间, 在保证计算结果精度的前提下, 实际计算时一个试件各列的N1i均取平均值, 即采用将管胞长度LT0的平均值代入(2)式中所算得的结果作为N1i的平均值。同样, 利用管胞并联纵向弹性模量公式(本研究理论模型中(21)式)进行计算的结果表明, 当NRT大于8 ×104后(相应计算涉及的管胞总数目为5 ×106~7 ×106), 在与测定结果一致的4位有效数字范围内几乎无变化, 故计算时的NRT值也使用由LR和LT的平均值代入(3)式中算出的结果。表 3列出了各类试件早晚材的N1i和NRT的平均值(按四舍五入法则以整数列出), 以及相应的晚材率和射线组织比量。
|
|

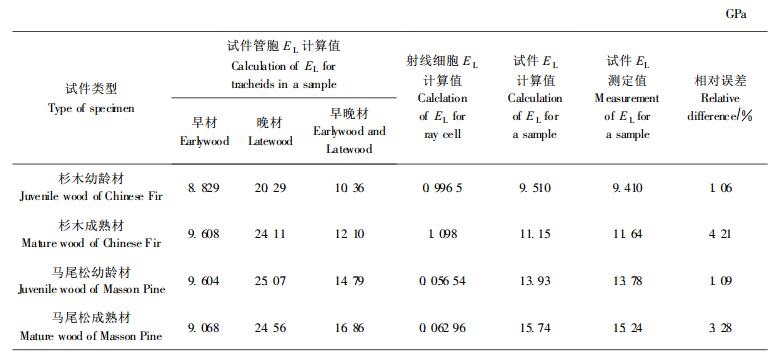
表 4列出了纵向弹性模量的有关计算结果, 其中所列4种类型的早材和晚材试件EL计算值, 是假定试件完全由早材管胞和晚材管胞构成, 由相应的管胞解剖结构参数以及表 2、3的数据, 根据本研究理论模型中的(4)、(9)、(10)、(16)、(21)式分别算出的管胞纵向弹性模量; 试件管胞早晚材EL的计算值, 是由其早材和晚材的EL利用该文中的(22)式计算所得; 试件EL的计算值, 则是由试件早晚材和木射线细胞的EL利用该文中的(23)式计算所得的有关数值; 相对误差以测定值为比较标准算出。
|
|
试件早材和晚材管胞EL的计算, 使用由Turbo C语言编制的程序在微型计算机上完成。正态分布随机数抽样子程序, 包括均匀分布随机数发生器、标准正态分布随机数抽样。均匀分布随机数发生器使用素数模拟式发生器, 在32位微机上其抽样序列周期不小于2.147 ×109(高惠璇, 1995), 大于1个试件可能包含的管胞最大的数目4.1 ×107, 从而保证管胞解剖结构参数计算中不出现重复抽样。同时, 采用抽样效率较高的“极坐标”抽样法实现标准正态分布的随机数的抽样(高惠璇, 1995), 均匀分布随机数发生器1次抽样, 可以获得2个服从标准正态分布的随机数。计算中所需管胞的5个解剖结构参数, 只需3次抽样即可完成。此外, 计算中管胞的搭接壁厚度取其径向和弦向壁厚度的平均值; 由于相邻的2个管胞共有1个胞壁, 计算时径向壁、弦向壁和搭接壁厚度取表 1所列值的一半。计算程序在CPU为P3、主频500MHz的微型计算机上进行, 根据试件早晚材管胞解剖结构的不同, 1个试件的早材或晚材纵向弹性模量的计算耗时为3~7 h左右。
3 结果与讨论根据表 4所列出的结果可知, 人工林杉木、马尾松幼龄材和成熟材4种试件的计算值与它们的测定值符合得十分好, 其相对误差最大为4.21%, 最小为1.06%, 平均为2.41%。这样的结果表明:首先, 本文关于管胞和射线细胞的结构模型虽然是一种简化的模型, 但能反映出针叶树木材细胞分子的基本特性, 与管胞和射线细胞的实际结构无大的偏离; 其次, 关于针叶树木材管胞纵向弹性模量的计算公式, 以及试件纵向弹性模量的计算方法, 比较准确地揭示了人工林杉木、马尾松的微观和宏观弹性特性。
从木材细胞的结构和弹性模量出发求得试件宏观弹性模量, 是本研究工作的基本精神与特征, 本文提出的模型与计算方法定量地描述了针叶树木材细胞与宏观纵向弹性模量的相关性, 即试件纵向弹性模量随木材细胞解剖结构变化的规律性, 可以用来研究和探索人工林杉木、马尾松及解剖结构与其类似的针叶树木材纵向弹性特性的一些变化规律。以下就人工林杉木、马尾松幼龄材和成熟材细胞解剖结构的变化, 对其宏观纵向弹性模量的影响进行一些讨论。射线细胞是影响木材材性的一个重要的因素(Schniewind, 1959; Burgent et al., 2001), 根据表 4中列出的计算结果可以发现, 射线的存在使试件纵向弹性模量减少, 这是因为射线的随机分布, 使得木材纵向的对称性与均匀性进一步破坏, 从而使木材纵向弹性模量变少。不过射线是树液及营养物质在生活中树木的径向通道, 在实际中难以改变它的存在。下面只针对管胞进行其解剖结构参数与试件纵向弹性模量相关性的讨论。
设想其它情况不变时, 4种试件中管胞的晚材率及长度、管胞的径向和弦向直径、径向和弦向直径胞壁厚度各自单独增加5%, 由此算出的试件纵向弹性模量在表 5中前4列中列出。表 5的最后一列, 则是晚材率及管胞的长度、径向和弦向直径、径向和弦向直径胞壁厚度同时增加5%时, 试件纵向弹性模量的计算值。经比较和分析后可以得到如下的结论。
|
|

由表 4可以看到, 4种试件晚材的纵向弹性模量是早材的2.3~2.7倍。由于两者之间存在着较大的差异, 在针叶树木材试件其它因素不变时, 其纵向弹性模量将以稍大于晚材率增幅而增大。例如当4种试件的晚材率较表 3所列的值增加5%, 即晚材率分别为18.40、22.20、38.52、53.40%时, 计算出的纵向弹性模量增加幅度分别为5.48、5.86、6.16、5.52%(见表 5), 均以大于相应晚材率的幅度增加。由此可知, 针叶树木材的晚材率对于其纵向弹性模量的影响大, 增加木材晚材率对于提高木材纵向弹性模量的效果明显。
3.2 管胞长度的影响如若管胞的其它因素不变, 对于一个尺寸确定的木材, 管胞长度的增加将引起木材纵向串联的管胞数目的减少, 这意味着试件纵向弹性模量计算式中求和的串联管胞数目减少, 从而使求和的结果变大。表 5中第3列的数字显示出了这样的结果, 管胞长度增加时木材的纵向弹性模量也增加, 但增加的平均幅度不大, 为1.72%。值得注意的是, 除人工林杉木成熟材外其它类型试件纵向弹性模量的增幅呈现出一个确定的趋势, 即管胞长度平均值大则其增幅大。
3.3 管胞直径的影响表 5中第5列所示的数字, 是其它因素不变时, 将管胞径、弦向直径同时增加5%后计算所得到的结果。由于管胞长度与胞壁厚度不变, 直径的增加则意味着管胞管状体的中空部分比例增大, 由管胞中空管状体纵向弹性模量的计算公式可知, 管胞的纵向弹性模量减少。同时, 直径的增加也使试件横截面所排列的管胞减少, 从而也导致试件纵向弹性模量减少。试件纵向弹性模量随管胞直径增大而减少的幅度比较大, 且管胞径、弦向直径平均值大者减少的幅度小, 而管胞长度平均值大者减少的幅度大。
3.4 管胞壁厚度的影响管胞壁厚度增加则木材的实质物质, 即MP层、3S层的物质增加了, 且空隙部分减少。空隙部分的弹性模量可视为空气的模量, 而常温下空气的弹性模量为105Pa数量级(内尔康, 1983), 较木材实质物质的弹性模量要小4~5个数量级, 这必然会使木材整体的纵向弹性模量增大。由表 5的计算结果可以看出, 管胞壁厚度变化所引起的4种类型试件纵向弹性模量平均变化幅度为5.85%, 较晚材率、管胞长度、管胞直径增大所引起的变化的幅度都要大, 这说明其对纵向弹性模量影响是很大的。还要说明的是, 其它因素不变时试件管胞壁厚度增加则其密度增大, 试件纵向弹性模量随管胞壁厚度增加而增加, 与木材的弹性模量随木材的密度增加而增加的结论一致(Bogig et al., 1982; 渡治人, 1984; Zhang, 1994; Leban et al., 1999)。
作为对比, 表中也列出了试件晚材率、管胞长度、管胞径向和弦向直径、胞壁径向和弦向厚度同时增大5%时, 试件纵向弹性模量的计算结果, 上述因素同时增加的结果是纵向弹性模量有一较小幅度的增加。通过观察和分析表 5中所有计算结果, 这里的增加幅度并不是前述4种变化的简单叠加, 而且尽管计算结果显示出与管胞直径增大时相反的变化趋势, 但不能就此推定这是一个确定的规律性。
本文使用的试件纵向弹性模量测定值, 是利用抗弯弹性模量测定结果所得出, 因而上面的讨论对于试件抗弯弹性模量也是适用的。
4 结论运用针叶树木材管胞和射线细胞的结构模型, 以及纵向弹性模量计算公式和方法, 计算了人工林杉木、马尾松幼龄材和成熟材试件纵向弹性模量, 其与常温条件下相应气干试件的测定值符合得十分好, 相对误差最大为4.21%, 最小为1.06%, 平均为2.41%。因此, 本研究所得的结果, 不仅建立了关于由木材微观的层次研究木材宏观弹性的理论途径, 而且提供了计算预测人工林杉木、马尾松及解剖结构类似的针叶树木材纵向弹性模量的实用方法。这一方面有助于人工林针叶树木材力学性质及其变化规律的理论研究, 另一方面也有助于开展人工林针叶树木材材性、材质的计算与预测。本文同时利用已经建立的模型、计算公式和方法, 计算预测了当晚材率、管胞长度、管胞径向和弦向直径、管胞壁径向和弦向厚度增加5%时, 4种人工林杉木、马尾松木材试件纵向弹性模量的变化情况。结果表明:试件的纵向弹性模量随晚材率、管胞长度、管胞壁厚度的增加而增加, 其中, 晚材率和管胞壁厚度增加引起的纵向弹性模量增加的幅度较大且两者较接近, 管胞长度增加所引起的纵向弹性模量增加的幅度较小; 试件的纵向弹性模量随管胞直径增加而减小, 其减小幅度较为明显。
纵向弹性模量计算预测的结果, 揭示了同一树种的试件中晚材率及管胞解剖结构引起纵向弹性模量变化的数量关系, 这为应用转基因、遗传育种等现代生物技术控制和改变针叶树木材的材质、材性, 提出了具体的实施方向和目标, 具有一定的应用价值。
北京林学院. 1983. 木材学. 北京: 林业出版社, 73-76.
|
渡治人[日]著, 张勤丽, 张齐生, 张彬渊译.木材应用基础.上海: 上海科学技术出版社, 1984: 374-375
|
高惠璇. 1995. 统计计算. 北京: 北京大学出版社, 98-101-146-148.
|
侯祝强, 姜笑梅, 殷亚方. 2001. 马尾松管胞长度分布规律及计算机随机模拟. 林业科学研究, 14(3): 271-277. DOI:10.3321/j.issn:1001-1498.2001.03.006 |
江泽慧, 费本华, 侯祝强, 等. 2002. 针叶树木材细胞力学及纵向弹性模量计算———纵向弹性模量的理论模型. 林业科学, 38(5): 101-107. DOI:10.3321/j.issn:1001-7488.2002.05.017 |
姜笑梅, 许明坤. 1994. 人工林和天然林杉木幼龄材和成熟材解剖性质的差异及其在径向变异上的比较研究. 世界林业研究, 7(专集): 1-13. |
姜笑梅. 1995. 人工林和天然林马尾松幼龄材和成熟材解剖性质比较研究. 世界林业研究, 8(专集): 1-9. |
骆秀琴, 管宁. 1995a. 人工林和天然林杉木幼龄材与成熟材力学性质的差异. 世界林业研究, 8(专集): 172-180. |
骆秀琴, 管宁. 1995b. 人工林和天然林马尾松幼龄材与成熟材力学性质的差异. 世界林业研究, 8(专集): 181-188. |
M内尔康[英]著, 倪锄非译.力学与物质的学性质.北京: 科学出版社, 1983: 199-201
|
中华人民共和国国家标准GB1927-43-91.木材力学性质试验方法.北京: 中国标准出版社, 1991: 38-41
|
Bodig J, Jayne B A. 1982. Mechanics of wood and wood composites. New York: Van Nostrand Reinhold Company Inc., 149-158: 488-491. |
Burgent I, Bernasconi A, Nikas K J, et al. 2001. The influence of rays on the transverse elastic anisotropy in green wood of deciduous tres. Holzforschung, 55(5): 449-454. |
Gillis P P. 1970. Elastic moduli for plane stress analysis of unidirectional composites with anisortopic rectangular reinforcement. Fiber Sci Tech, 2: 193-210. DOI:10.1016/0015-0568(70)90002-3 |
Leban J M, Haines D W. 1999. The modulus of elasticity of hybrid larch predicted by density, rings per centimeter, and age. Wood Fiber Sci, 31(4): 394-402. |
Mark R E, Gillis P P. 1970. New models in cell-wall mechanics. Wood and Fiber, 2(2): 79-95. |
Mark R E. 1967. Cell wall mechanics of tracheids. Yale University Press, 233-234.
|
Schniewind A P. 1959. Transverse anisotropy of wood: A function of gross anatomic structures. For Prod J, 9(10): 350-359. |
Vysotskaya E, Vaganov A. 1989. Components of the variablility of radial cell size in tree rings of conifers. IAWA Bulletin N S, 10(4): 417-428. DOI:10.1163/22941932-90001134 |
Watanabe U. 1998. Shrinking and elastic properties of coniferous wood in relation to cellular structure. Wood Research, 85: 1-47. |
Zhang S Y. 1994. Mechanical properties in relation to specific gravity in 342 Chinese woods. Wood Fiber Sci, 26(4): 512-526. |