 2003, Vol. 39
2003, Vol. 39文章信息
- 谢芳.
- Xie Fang.
- 乳源木莲优质干材混交培育的研究
- CULTIVATING HIGH-QUALITY STEMWOOD OF MANGLIETIA YUYUANENSIS BY MIXING WITH CUNNINGHAMIA LANCEOLATA
- 林业科学, 2003, 39(2): 84-90.
- Scientia Silvae Sinicae, 2003, 39(2): 84-90.
-
文章历史
- 收稿日期:2001-10-15
-
作者相关文章


保护和发展阔叶树资源, 鼓励阔叶树人工造林并实行优惠政策是许多林业先进国家作为保护生物多样性、防治地力衰退和提高林产品附加值的重要途径。如英国对橡树等阔叶树种的造林补助金高于针叶树的1.5倍。我国台湾从20世纪70年代就开始注重珍稀及有潜力树种的培育, 重点开展牛樟等5大珍贵阔叶树的选育和培育研究。目前我国木材消费结构发生了巨大变化, 大径材和珍贵阔叶用材十分短缺。福建省是我国的重要商品材产区, 从20世纪80年代初开始, 在大力发展杉木商品林的同时, 就已意识到培植阔叶林资源对林业可持续经营的重要性, 积极发掘和推出了一批优良的乡土阔叶用材树种, 大规模地营造各种混交模式的针阔混交林, 成效显著(刘春华等, 1993; 庄孟能等, 1994; 王忠平等, 1996;范辉华, 2001)。然而当初营造混交林的目的多着眼于提高和维护杉木人工林地力, 而较少考虑优质阔叶用材的培育。
乳源木莲(M.yuyuanensis Law)是新近发掘的木兰科优良阔叶树种, 自然分布于浙江、安徽、江西、福建、湖南、广东等省, 生长迅速, 树干通直, 材质优异, 适应性和耐寒力强, 是与松杉混交的理想树种(王忠平等, 1996)。本文将用营造的14 a生和16 a生杉莲混交林, 以杉木(C.lanceolata(Lamb.)Hook)作为伴生树种, 从针阔混交培育优质干材的角度, 研究造林立地环境和混交措施对乳源木莲干材生长、干形培育和材质材性的影响, 为速生优质阔叶木的培育提出适宜于人工造林的经营模式与技术措施。
1 材料与方法试验数据采集自福建省沙县林业局1986和1988年营造的15 hm2杉莲混交林。沙县地处福建省中西部(117°49'E, 26°26'N), 属中亚热海洋性季风气候, 年均温19.2 ℃, 1月均温8.7 ℃, ≥ 10 ℃的积温6 135 ℃, 无霜期291 d, 年降雨量1 618 mm。自1984年福建省就开始乳源木莲的采种育苗, 大力营造杉莲混交林。杉莲混交林的立地为低山丘陵, 立地条件较好, 土壤深厚, 为砂岩、花岗岩等发育的山地红壤和黄红壤。造林地前茬为天然林, 部分是杉木采伐迹地和撩荒地。目前, 乳源木莲与杉木的混交效果显著, 乳源木莲生长远优于杉木, 适宜以乳源木莲优质干材培育为经营目标。
2001-07, 按不同造林年份、不同生育地环境(海拔、坡向、坡度、坡位)和不同混交比例, 在沙县富口银地(1986年造林)和水南林场(1988年造林)的杉莲混交林中设立21块调查样地, 样地大小为20 m ×20 m, 各调查样地乳源木莲的立地环境和重要影响因子见表 1。
|
|
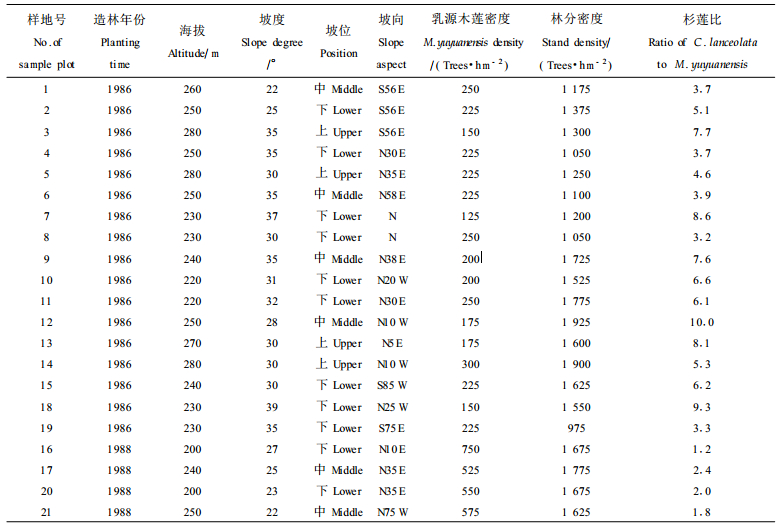
对样地内的所有乳源木莲进行每木调查, 测定指标包括树高、胸径、冠幅、枝下高、树干通直度(按5分制统计, 分数越高越通直)、下部3m段树干的圆满度(用3 m高处直径与胸径之比表示)、侧枝形成分叉干特性、分枝性状(1级活枝数、最大侧枝基径、长度和枝角)等指标, 并在每木胸高处的上坡方位用6 mm直径的生长锥取得由髓心至树皮的无疵木芯, 测定其木材基本密度。对于标准地内的杉木, 测定每木胸径和平均木的树高。
选用1986年造林的17个调查样地乳源木莲的测定数据, 首先从造林成活率、干材高径生长、侧枝形成分叉干率、干形、分枝特性和材质材性等对混交培育乳源木莲优质干材进行总体评价, 然后通过方差分析以验证造林地环境和林分密度等重要影响因子对乳源木莲干材培育的作用。由于各调查样地海拔差异较小, 这里重点就坡度、坡位、坡向、乳源木莲密度、林分密度和杉莲比6个影响因子进行分析, 6个因子具体分级见表 2。以样地内乳源木莲单株测定值为单位进行方差分析。因试验数据不平衡, 方差分析均采用SAS/STAT软件中的GLM程序。在方差分析时树干通直度、1级活枝数和最大侧枝角数据经X-1/2转换。
|
|
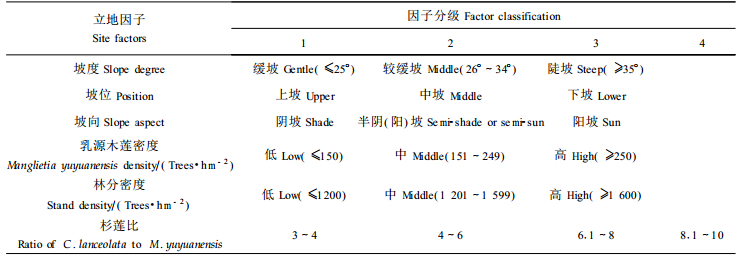
乳源木莲虽是木兰科的常绿树种, 但经过多年的育苗试验, 已完全掌握根系发达之优质苗木的培育技术, 因此只要把握好造林季节以及对苗木适宜的修剪等产后处理, 其栽植成活率不低于混交树种杉木, 达95%以上。试验调查未发现乳源木莲植株有枯死现象, 概因其有一定的耐荫性, 与杉木种间关系协调之故。
与杉木生长习性不同, 乳源木莲早期生长较缓, 造林后头几年的高径生长远低于混交树种杉木, 10 a生左右才进入其干材速生期。图 1显示, 在混交造林后第8 a, 乳源木莲的高径生长还低于杉木, 9 a生时其高生长超过了杉木, 胸径与杉木接近。在本次调查的16 a生时, 乳源木莲的干材生长远超过杉木, 其平均树高为12.9 m(11.5~15.4 m), 平均胸径为21.8 cm(18.7~28.0 cm), 分别大于杉木高径的15%和73%, 而乳源木莲单株材积接近杉木的3倍, 这充分说明, 通过与杉木的混交造林, 可以达到培育大径阶的乳源木莲优质干材的目标。

|
图 1 以杉木为参照, 混交乳源木莲之干材树高(左)、胸径(右)生长及年龄效应 Fig. 1 Height (left) and DBH (right)growth of stemwood and its age effect in mixed M.yuyuanensis stand, comparing with C.lanceolata ■乳源木莲Manglietia yuyuanensis; □杉木Cunninghamia lanceolata. |
侧枝形成分叉干是许多阔叶树和一些针叶树固有的遗传特性。如马褂木一般形成2~3个分叉干, 多的达5个分叉干, 不同种源的分叉干率差异很大, 在26.35%~73.76%(李建民, 2001; 詹明勋等, 1998)。台湾红桧的分叉干率达30%, 分叉部位发生在0~0.5 m处最多(李建民等, 2001)。按调查样地所有植株统计, 乳源木莲侧枝形成分叉干率达41.42%, 已超过马褂木种源平均值(表 3)。在有分叉的乳源木莲植株中以形成一个分叉干为主, 而2叉干和多叉干较少。与马褂木和台湾红桧不同的是, 乳源木莲的分叉干形成多发生在植株较高部位(大于50 cm高处)。根据乳源木莲侧枝分叉干形成的规律, 不仅要求进行适时的除萌, 而且要求修剪才能培育速生优质之干材, 仅依靠混交单项措施还不够。
|
|

阔叶木优质干材培育一般集中在树干下部3 m段, 要求通直圆满、节小节少。调查统计发现(表 4), 16 a生的乳源木莲平均枝下高为2.9 m, 已基本满足3 m段干材培育的要求, 但不同样地乳源木莲的枝下高差异较大, 变化在2.0~5.0 m之间。乳源木莲的干形在样地间变异较小, 较少受环境影响。乳源木莲最大侧枝长为3.8 m, 最大侧枝基径为4.9 cm。木材测定发现, 乳源木莲木材白度较高, 基本密度中等(0.36 g·cm-3), 与引种四川桤木(0.35 g·cm-3)相近(陈炳星等, 2000)。乳源木莲其它的一些分枝性状详见表 4。
|
|
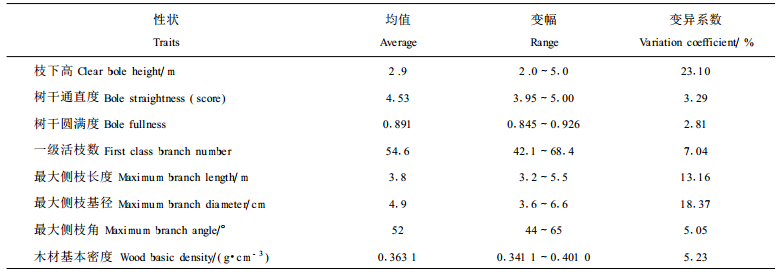
从乳源木莲干材生长与其干形、分枝和木材密度的相关分析结果第2期谢芳:乳源木莲优质干材混交培育的研究87来看(表 5), 干材的高径生长与树干通直度、最大侧枝角相互独立。不难理解, 随着高径生长的提高和冠幅的拓展, 一级活枝数增多, 侧枝加长, 侧枝基径增粗。研究发现, 树干圆满度和枝下高有随干材高生长增加而增大和升高的趋势。乳源木莲木材基本密度与高径生长仅呈弱度负相关关系。
|
|
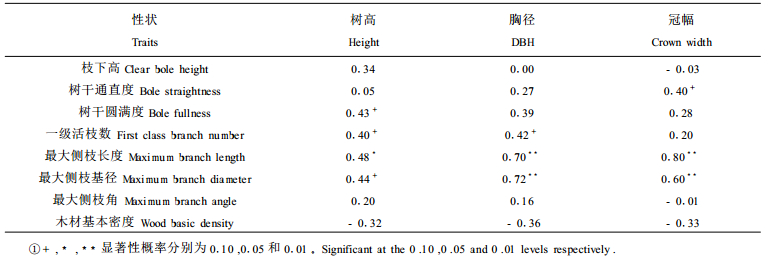
立地环境涉及混交林所处的海拔高度、坡度、坡位、坡向, 以及由此导致的水、温、肥和光照等微环境的改变。由于杉莲混交林多为低丘山地, 海拔高度大致在200~300 m, 各调查样地的海拔高度差异很小, 因此本文重点对坡度、坡位和坡向等立地影响因子进行分析, 其方差分析结果及性状均值分别见表 6和表 7。
|
|

|
|
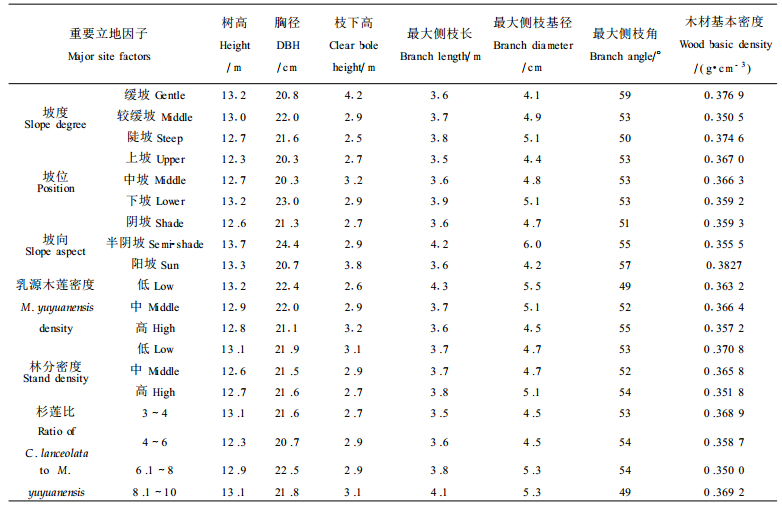
乳源木莲的干形在样地间不存在显著差异, 较少受造林立地微环境的影响, 坡度、坡位和坡向效应不显著。研究发现, 乳源木莲一级活枝数似是一个独立的分枝性状, 与造林立地环境无关。而最大侧枝长和侧枝基径较易受微环境条件影响。在本试样调查期, 虽然乳源木莲已处在干材速生增长期, 但仍具有并且也需要有一定的耐荫性。半阴(阳)坡或下坡恰好符合乳源木莲干材速生期的这种生物学习性, 树高和胸径生长量较大, 并随着树冠的拓展, 侧枝增长变粗。对于最大侧枝角, 这里仅发现存在一定的坡度效应, 生长在缓坡的乳源木莲其侧枝较平展。上面已述及, 乳源木莲的干材生长与木材基本密度仅呈微弱的负相关, 这可解释为什么木材基本密度存在一定的坡度和坡向效应, 但却无一定规律性。

|
图 2 不同坡度的干材树高生长(左)和枝下高(右) Fig. 2 Tree height (left) and clear bole height (right)of M.yuyuanensis planted in different slopes |
1986年营造近7 hm2杉莲混交林大多采用同一混交模式(杉莲比为3:1, 行间混交)和同一造林密度。因经过多次间伐, 各样地乳源木莲和杉木(或林分)的保留密度和杉莲比已完全不一样。方差分析结果显示(表 6), 乳源木莲现有密度对干材基本密度已产生显著影响, 对干材其它性状的作用较小。当乳源木莲保留较低密度时, 树高和胸径生长加速, 侧枝有增长加粗的趋势(虽然未达显著水平), 此时干材基本密度将提高0.06 g·cm-3, 这概缘因于晚材比例增加之故。研究发现, 现有林分密度对乳源木莲的树高生长、侧枝长和基径有显著作用。
研究混交比例对乳源木莲优质干材培育的影响对于确定适宜的混交方式, 以及进行适时间伐具有重要意义。根据1986年造林的17个调查样地的统计分析结果, 杉莲比对乳源木莲干材生长似无影响, 而对枝下高和侧枝性状有明显的作用。随着杉莲比的降低, 干材枝下高将减小。然而根据1988年造林, 立地环境完全一致的两个样地测定结果则得出相反的结论, 即杉莲比的降低将使乳源木莲的干材枝下高升高, 杉莲比为1:1时乳源木莲枝下高为4.1 m, 而当杉莲比为2:1时, 枝下高仅为3.0 m(表 8)。这里出现两种相反的结论其实不难解释。对于1986年营造林的杉莲混交林, 虽然各样地的杉莲比差异较大, 但单位面积内乳源木莲的保留数株都处在一个较低水平(150~300株·hm-2)。如此, 乳源木莲间难以通过竞争达到自然整枝, 而随着杉莲比的提高, 由于杉木株数的增多而使乳源木莲受到强烈的生长竞争, 结果乳源木莲的枝下高升高了。对于1988年营造的杉莲混交林, 则是另一种情形。由于乳源木莲植株数较多(525~750株·hm-2), 乳源木莲间竞争强烈, 因杉莲比的降低, 这种竞争尤为强烈, 结果乳源木莲的枝下高将增高。
|
|

研究揭示, 乳源木莲早期生长缓于杉木, 10 a生左右进入干材速生期, 16 a生时其单株材积几乎是杉木的3倍。乳源木莲树干通直、圆满, 较少受立地环境和育林措施影响, 但其侧枝形成分叉干特性显著而影响其优质干材的培育。乳源木莲侧枝形成分叉干率达41.42%, 以形成一个分叉干、发生在较高部位为主, 应进行适时的除萌修枝方可保证高质量干材的培育。乳源木莲木材密度中等, 超过同龄的杉木, 适宜以胶合板材和实木用材为培育目标。
在低山丘陵区, 坡度是影响乳源木莲干材生长的主要立地因子, 而坡位和坡向作用较小。不同坡度反映的是立地条件的差异。坡度平缓意味着立地质量高, 结果乳源木莲干材生长量大, 枝下高长。
适宜的造林密度和混交比例有利于培育大径阶的优质干材。1986年营造的杉莲混交林虽采用行间混交, 杉莲比为3:1的同一混交模式, 但由于对杉木和乳源木莲实施了多次间伐, 各样地保留密度和杉莲比相差很大, 但总的来说, 单位面积的乳源木莲株数相对较少(150~300株·hm-2)。在这种情况下, 乳源木莲干材生长量很大, 杉莲比对其干材生长似无影响, 而对枝下高和侧枝性状有明显的作用。由于单位面积乳源木莲株数较少, 无法通过乳源木莲间竞争, 而只通过与杉木的竞争达到自然整枝, 杉莲比低意味着杉木株数的减少和植株间竞争的降低, 这样干材枝下高将减小。而对1988年营造林的杉莲混交林, 由于单位面积乳源木莲的株数很多, 虽然乳源木莲干材的高生长不受影响, 但与1986年造的乳源木莲相比, 径生长小了很多, 随着杉莲比的降低, 乳源木莲间植株将发生强烈竞争, 结果使枝下高升高。根据上述林分密度和杉莲比之于乳源木莲干材培育的作用机理, 为达到培育枝下高长、径阶大的干材培育目标, 首先应适当降低混交林的初植密度, 尤其要降低单位面积乳源木莲的株数, 以促进乳源木莲干材的高径生长; 其次是杉莲混交比不应过低, 宜采用3:1或更高的杉莲比进行混交并适时间伐, 虽不利于乳源木莲枝下高的增长, 但可增加乳源木莲生长的营养空间, 以利其干材的快速增粗。
陈炳星, 李光荣, 周志春, 等. 2000. 福建引种区四川桤木的木材化学组分分析与评价. 中国造纸, 19(4): 20-23. DOI:10.3969/j.issn.0254-508X.2000.04.004 |
范辉华. 2001. 杉木、拟赤杨混交对杉木持续生长的影响. 林业科学研究, 14(4): 455-458. DOI:10.3321/j.issn:1001-1498.2001.04.018 |
李建民. 2001. 马褂木地理遗传变异和优良种源选择. 林业科学, 37(4): 41-49. DOI:10.3321/j.issn:1001-7488.2001.04.007 |
李建民, 谢芳, 封建文, 等. 2001. 北美鹅掌楸种源在福建省栽植之表现. 南京林业大学学报, 25(4): 26-30. DOI:10.3969/j.issn.1000-2006.2001.04.006 |
刘春华, 张春能, 郑燕明. 1993. 观光木及其混交林生态系统生物量和生产力研究. 福建林学院学报, 13(3): 267-272. |
王忠平, 陈水龙, 赖培淼, 等. 1996. 乳源木莲造林试验研究初报. 福建林学院学报, 16(4): 315-318. |
詹明勋, 王亚男, 姜家华, 等. 1998. 重要生育地因子影响红桧造林木侧枝形成分叉干之研究. 中华林学季刊, 31(3): 239-250. |
庄孟能, 叶章善, 马祥庆. 1994. 杉木拟赤杨林分结构和生产力. 福建林学院学报, 14(4): 339-343. |