 2003, Vol. 39
2003, Vol. 39文章信息
- 孙志虎, 王庆成.
- Sun Zhihu, Wang Qingcheng.
- 应用PV技术对北方4种阔叶树抗旱性的研究
- THE DROUGHT RESISTANCE OF FOUR BROAD-LEAVED SPECIES IN THE NORTH OF CHINA WITH PV TECHNIQUE
- 林业科学, 2003, 39(2): 33-38.
- Scientia Silvae Sinicae, 2003, 39(2): 33-38.
-
文章历史
- 收稿日期:2002-09-23
-
作者相关文章



水分是树木生长的重要生态因子之一, 影响树木的生长发育及地理分布范围。世界上约有1/3的地区, 我国约有52%国土面积属于干旱、半干旱地区(郭连生等, 1998)。土壤水分条件严重制约造林绿化工作。研究树种抗旱能力是实现干旱、半干旱地区造林绿化的重要前提。茶条槭、山荆子、山桃和山梨是东北林区常见的树种, 树形美观、季相明显, 具有抗寒、抗风雪、抗烟害等多种优点, 具极高的绿化潜力(周以良等, 1986)。关于山桃抗旱性的评价有过一些研究(李吉跃等, 1993; 李海涛等, 1998; 郭连生等, 1998; 李良厚等, 1999), 其它3树种抗旱性的研究却未见报道。借助PV曲线技术, 可计算出被测植物体的多项水分状况参数(Schulte et al., 1985; Grossnickle, 1989; Zine et al., 1993)。越来越多研究者应用由PV曲线所确定的植物水分状况参数(指标)来评价植物种的耐旱特征并比较不同植物种的抗旱性强弱(张建国等, 1994; 李吉跃等, 1994; Boucher et al., 1995; White et al., 1996; 杨敏生等, 1997; Jordi et al., 1997; John et al., 1999; 2001;Kurt et al., 1999; Maria et al., 2001; Donald et al., 2001)。本文应用PV技术确定茶条槭、山荆子、山桃和山梨水分参数, 比较它们的抗旱特征及抗旱性强弱, 旨在为选择抗旱树种, 提高绿化造林效率提供科学依据。
1 材料和方法试验在东北林业大学尖砬沟森林培育实验站温室内进行。选取茶条槭(Acer ginnala)、山荆子(Malus baccata)、山桃(Prunus davidiana)、山梨(Pyrus ussuriensis)当年生播种苗为试验材料, 其平均苗高、地径依次为17.3 cm、2.20 mm, 20.8 cm、3.01 mm, 21.4 cm、2.99 mm, 13.4 cm、3.83 mm。采用盆栽控水法进行不同程度的水分处理(毛达如, 1994)。栽植桶高25 cm, 上口直径30 cm, 下底直径23 cm。桶内装入等量土壤并适当拍实。装土后, 随机用环刀取土样, 测定土壤水分-物理性质(刘光崧, 1996), 重复10次。测得基质容重0.76 g·cm-3, 饱和含水量86.9%, 毛管持水量81.1%, 最小持水量50.1%, 非毛管孔隙4.4%, 毛管孔隙61.7%, 总孔隙度66.1%。每桶栽植苗木4株。苗木正常供水生长4周后, 将每个树种苗木分成4组, 每组5盆, 每组为一个控水等级。用称重法进行控水处理, 各处理的土壤相对含水量分别为, A:75%;B:61%;C:46%;D:35%。每天下午5:00称重, 补充失去的水分。
土壤形成相应水分梯度4周后, 从供试苗木中截取10 cm左右长的枝条, 立即用万分之一电子天平称其鲜重, 然后将小枝插入盛有清水的烧杯中, 置于阴暗高湿条件下, 饱和吸水24 h, 当小枝达到饱和状态时取出, 迅速称饱和鲜重并立即装入压力室(柴宝峰等, 2000)。本试验采用Hammel法(王万里, 1984), 分次测定压出的水量, 最后取出样品时再测定鲜重, 并于75℃下烘干至恒重, 然后计算出全过程中样品的相对水分亏缺和渗透水含量。以依次测得的各次平衡压的倒数为纵坐标, 相应的相对水分亏缺(RWD)为横坐标, 绘制PV曲线。借助PV曲线计算出每个供试小枝的有关水分参数。由于PV曲线重复性好(李吉跃, 1989; 张建国等, 1994), 每个处理仅测1次PV曲线。
2 结果与讨论 2.1 不同土壤水分对叶水势的影响4树种苗木清晨叶水势在所设计的土壤水分梯度内均较高, 随土壤含水量的降低呈减小趋势(表 1)。A处理条件下, 4树种苗木叶水势(ψw), 均在-0.15 MPa左右; D处理条件下, 山梨水势最低且降低幅度最大, 其它3树种水势较高, 降低幅度较小。随着土壤含水量的降低, 植物降低其叶水势, 有利于苗木从土壤中吸收水分(阮成江等, 2000)。表明山梨吸收水分能力强。
虽然4种处理间土壤相对含水量相差最大达40%, 但不同处理叶水势均较高, 其原因可能是清晨时, 苗木通过气孔从温室内湿度较大的空气中吸入了较多的水分(Boucher et al., 1995)。
2.2 不同土壤水分对束缚水的影响随着土壤含水量的变化, 4树种苗木的束缚水含量表现出不同的变化趋势(表 1)。山荆子苗木束缚水含量(Va)随土壤含水量的降低呈降低趋势。在D处理条件下, 山荆子苗木束缚水含量较A处理条件下降低31.4% (表 1)。束缚水含量降低, 由束缚水所引起的渗透势升高, 而山荆子在D处理条件下叶水势降低, 表明山荆子具有强的渗透调节能力。渗透调节在树木体内可通过3条途径来实现:束缚水含量增加; 细胞体积减小; 细胞内溶质增加(Wilson, 1980; 汤章城, 1983)。在一定条件下树木会以其中之一为主。由此可见, 在干旱胁迫条件下, 山荆子是通过细胞体积减小或增加溶质含量来实现渗透调节。茶条槭、山梨和山桃束缚水含量在胁迫条件下出现不同程度的升高(表 1)。山桃、茶条槭在土壤含水量降至B处理后, 其束缚水含量就趋于稳定, 表明在C、D处理条件下, 由束缚水含量所引起的渗透调节作用消失; 山梨由束缚水含量引起的渗透调节作用维持至C处理。这不仅表明3树种能通过提高束缚水比例来实现渗透调节, 而且反映3树种由束缚水含量所引起的渗透调节作用是有一定限度的。从束缚水含量上升的幅度看, 山梨渗透调节能力最大(32.25%), 山桃其次(27.64%), 茶条槭最小(16.21%)。
|
|
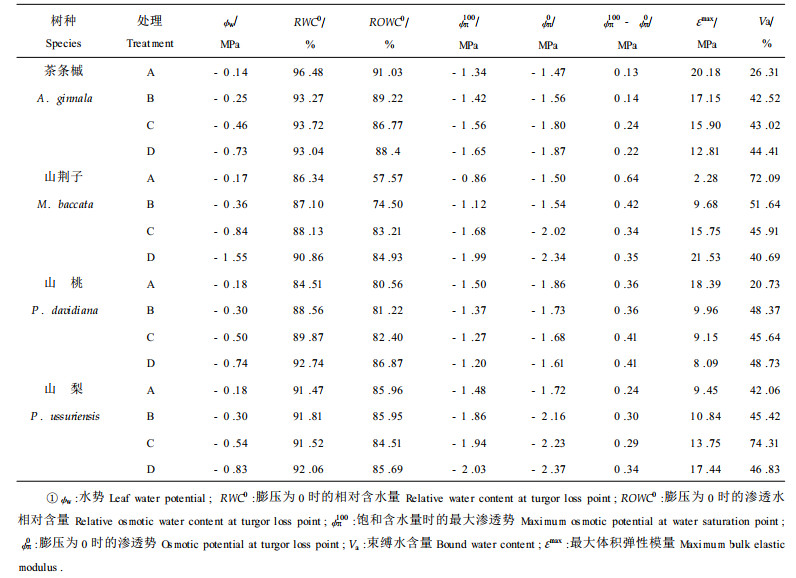
随着土壤含水量的降低, 4树种苗木饱和含水时的最大渗透势(ψπ100)的变化趋势不同(表 1)。山桃苗木随着土壤含水量的降低, ψπ100值升高(表 1), 其它3树种ψπ100随土壤含水量的降低呈降低趋势:山荆子降低幅度最大(131.4%), 山梨次之(37.2%), 茶条槭最小(23.1%)。在D处理条件下, ψπ100由小到大依次为山梨、山荆子、茶条槭、山桃, 反映在由ψπ100所指示的抗旱能力上, 由强至弱依次为山梨、山荆子、茶条槭、山桃。
在干旱胁迫条件下, 4树种苗木膨压为0时的渗透势(ψπ0)的变化趋势和ψπ100的变化趋势基本相同(表 1)。随土壤含水量的降低, 山桃ψπ0升高, 由ψπ0所反映的细胞维持膨压的能力降低。茶条槭、山荆子、山梨ψπ0值降低, 降低幅度依次为山荆子(56%)、山梨(37.8%)、茶条槭(27.2%), 表明3树种随土壤含水量的降低, 由ψπ0所反映的细胞维持膨压的能力升高。在D处理条件下, ψπ0由小到大依次为山梨、山荆子、茶条槭、山桃, 反映在由ψπ0所指示的抗旱能力上, 由强至弱依次为山梨、山荆子、茶条槭、山桃。
随土壤含水量的减少, 茶条槭、山桃、山梨ψπ100和ψπ0差值增加, 表明3树种在干旱胁迫下, 叶片渗透调节能力得到一定程度提高; 山荆子虽然随着土壤含水量的降低, ψπ100和ψπ0差值减小, 但ψπ100和ψπ0的下降幅度最大, 分别下降1.13 MPa和0.84 MPa, 表明山荆子具有极强的渗透调节能力。根据ψπ0值及ψπ100和ψπ0差值可判断苗木渗透调节能力:山荆子和山桃渗透调节能力最强, 山梨次之, 茶条槭最弱。
2.4 不同土壤水分对初始质壁分离时的相对含水量和相对渗透水含量的影响由表 1可见, 4树种在D处理条件下叶片组织细胞初始质壁分离时的相对渗透水含量(ROWC0)均在80%以上。这同Cheung等(1975)研究银杏、杨树等的结果相一致, 认为这应归功于由细胞壁特性所决定的“渗透调节”能力。4树种苗木ROWC0均在80%以上, 表明4树种由细胞壁特性所决定的“渗透调节”能力均很强。随着土壤含水量的降低, 茶条槭的叶片组织细胞初始质壁分离时的相对含水量(RWC0)和相对渗透水含量(ROWC0)呈下降趋势, 表明随着土壤干旱胁迫的发展, 茶条槭忍耐脱水能力得到提高, 由细胞壁特性所反映的“渗透调节”能力减弱。茶条槭RWC0和ROWC0值在各干旱胁迫条件下均较高; 山荆子、山桃RWC0和ROWC0随土壤含水量的降低呈升高趋势, 反映山荆子、山桃原生质耐脱水能力对干旱胁迫的适应性差, 由细胞壁特性所反映的“渗透调节”能力经过水分胁迫处理得到提高。山荆子在各干旱程度上RWC0和ROWC0值均较低, 表明山荆子在原生质忍耐脱水方面较其它树种占有很大优势。山梨RWC0和ROWC0值基本保持不变。根据各干旱程度RWC0和ROWC0值及其变化趋势, 可判断4树种原生质忍耐脱水能力:山荆子细胞忍耐脱水能力最强, 山桃、山梨次之, 茶条槭最差。
2.5 不同土壤水分对组织细胞最大弹性模量的影响由表 1可知, 4树种之间, 组织细胞最大弹性模量(εmax)存在明显差异。在A处理条件下, εmax由大到小依次为茶条槭、山桃、山梨、山荆子, 表明山荆子细胞最富有弹性, 维持膨压能力最强, 而山桃、茶条槭, 尤其是茶条槭, 细胞弹性较差, 维持膨压能力最弱。当苗木受到干旱胁迫时, εmax值发生了不同的变化。山梨、山荆子, εmax值表现不同程度的升高:在D处理条件下, 山荆子上升9.44倍, 山梨上升1.85倍。山梨、山荆子在遇到干旱胁迫时细胞弹性降低, 由细胞壁弹性所反映的维持膨压能力降低, 说明山梨、山荆子在干旱胁迫下主要靠渗透调节来维持膨压。山桃、茶条槭苗木受到干旱胁迫时, εmax值呈降低趋势, 表明干旱胁迫不仅没有使细胞壁变硬, 反而增强了细胞弹性, 山桃、茶条槭能通过改变细胞壁弹性来保持膨压, 这是适应干旱条件的特殊反应, 对提高其耐旱力具有积极作用。在D处理条件下, εmax由大到小依次为山荆子、山梨、茶条槭、山桃, 反映在细胞壁弹性上由强至弱依次为山桃、茶条槭、山梨、山荆子。
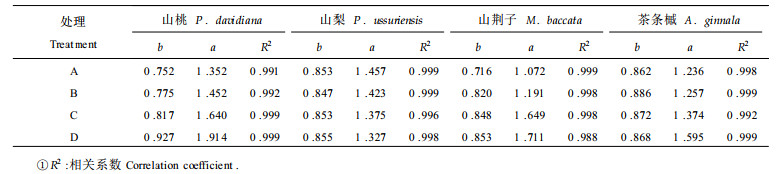
2.6 不同土壤水分对叶水势与膨压关系的影响不同土壤水分条件下, 4树种苗木膨压(ψp)与叶水势(ψw)均呈极显著直线相关关系, 相关系数均在0.99以上, 这与许多其它研究结果是一致的(Jones, 1981; 李吉跃等, 1994; 杨敏生等, 1997)。直线方程(ψp=a+bψw)中, a值表示苗木充分膨胀时叶片细胞所能达到的最大膨压; b值表示随叶水势下降, 膨压下降的速度, 能反映植物保持膨压能力的大小(李吉跃等, 1994)。在A处理条件下, 4树种a值:山梨>山桃>茶条槭>山荆子, 随土壤含水量的降低, 山桃、山荆子、茶条槭a值明显升高(表 2), 表明经过干旱胁迫处理苗木保持最大膨压能力得到加强; 山梨a值呈降低趋势, 细胞维持最大膨压能力减弱。在A处理条件下, 4树种b值:山荆子 < 山桃 < 山梨 < 茶条槭, 山荆子保持膨压能力明显高于其它树种。在干旱胁迫下, 树种b值变化趋势不同:山桃、山荆子b值明显升高, 保持膨压能力显著降低; 山梨、茶条槭膨压保持能力基本不变。在D处理条件下, 4树种b值:山荆子 < 山梨 < 茶条槭 < 山桃, 说明在干旱条件下, 膨压维持能力由强至弱依次为山荆子、山梨、茶条槭、山桃。
|
|
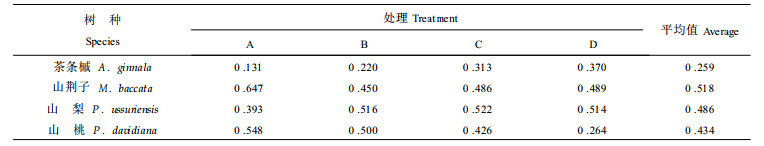
苗木的耐旱性受到多个水分参数的影响, 用不同的水分参数对其进行评价得出的结论不同(李良厚等, 1999)。为了作出准确的评价, 需要从多角度, 采用多个指标, 综合评价(刘友良, 1992)。需要说明的是, 由于植物的渗透调节能力都有一定限度(Kozlowski, 1983; Jones, 1981), 上述各水分参数的变化趋势, 仅是在试验范围内的结论, 当水分胁迫超过一定限度时, 其变化趋势可能发生改变。为了对树种耐旱性进行准确比较, 选用ψπ0、ψπ100、RWC0、ROWC0、εmax及膨压与叶水势直线方程中的b值作为参评指标, 将这6个指标分别在不同水分胁迫强度范围内进行比较, 综合评定。评定方法采用模糊数学反隶属函数进行定量转换(张建国等, 1994; 杨敏生等, 1997), 计算公式为:Uijk=1- (Xijk-Xkmin) / (Xkmax-Xkmin), 式中, Uijk为第i树种第j个水分胁迫强度对于第k项指标的隶属函数值; Xijk为第i个树种第j个水分胁迫强度第k个指标测定值; Xkmin、Xkmax为4树种第k项指标的最小值和最大值。Uijk值越大, 耐旱性越强。
从表 3可见, 随着土壤水分的减少, 茶条槭、山梨耐旱性增强; 山桃耐旱性降低; 山荆子在B处理条件下耐旱性最弱, 其耐旱性在干旱胁迫条件下, 有减弱的趋势。根据4树种不同土壤含水量下的综合评判结果, 4树种耐旱性由强至弱依次为山荆子、山梨、山桃、茶条槭。
|
|
4树种苗木在所设计的水分梯度内, 随土壤含水量的减少, 均能在一定程度上提高渗透调节能力, 但其实现渗透调节方式不同。在干旱胁迫条件下, 茶条槭、山梨、山桃能通过增加束缚水含量来实现渗透调节, 但其依靠改变束缚水含量来实现渗透调节作用有一定限度。山桃、茶条槭在土壤含水量降至B处理, 山梨在土壤含水量降至C处理后, 由束缚水含量所引起的渗透调节作用消失; 山荆子在干旱胁迫条件下通过细胞体积减小或增加溶质含量来实现渗透调节。综合来看, 4树种苗木渗透调节能力由强至弱依次为山荆子、山桃、山梨、茶条槭。
4树种忍耐脱水能力不同。山荆子细胞忍耐脱水能力最强, 山桃、山梨次之, 茶条槭忍耐脱水能力最差。在干旱胁迫条件下, 茶条槭能通过提高原生质忍耐脱水能力来适应干旱。山荆子、山桃原生质耐脱水能力对干旱胁迫的适应性差, 在干旱胁迫条件下其耐脱水能力减弱。
4树种维持膨压能力不同。山梨、山桃、山荆子维持膨压能力最强, 茶条槭维持膨压能力较差。在干旱胁迫条件下, 山桃、茶条槭通过改变细胞壁弹性来保持膨压; 山梨、山荆子在干旱胁迫下主要靠渗透调节来维持膨压。干旱胁迫能加强山桃、山荆子、茶条槭保持最大膨压的能力, 而降低山梨维持最大膨压的能力。
4树种耐旱性不同。耐旱性由强至弱依次为山荆子、山梨、山桃、茶条槭。由于树木耐旱能力的强弱不是一个绝对概念, 其排序只是相对而言。大量的研究表明, 山桃属强耐旱性树种(李吉跃等, 1993; 李海涛等, 1998), 本次试验表明山荆子、山梨的耐旱能力强于山桃, 综合来看山荆子、山梨亦属强耐旱树种, 茶条槭耐旱能力相对较弱。
柴宝峰, 李洪建, 王孟本. 2000. 晋西黄土丘陵区若干树种水分生理及抗旱性量化研究. 植物研究, 20(1): 79-85. |
郭连生, 田有亮. 1998. 运用PV技术对华北常见造林树种耐旱性评价的研究. 内蒙古林学院学报, 20(3): 1-8. |
李海涛, 陈灵芝. 1998. 暖温带森林生态系统主要树种若干水分参数的季节变化. 植物生态学报, 22(3): 202-213. |
李吉跃, 张建国, 姜金璞. 1994. 京西山区人工林水分参数的研究(Ⅱ). 北京林业大学学报, 16(2): 1-8. |
李吉跃, 张建国. 1993. 北方主要造林树种耐旱机理及其分类模型的研究(Ⅰ). 北京林业大学学报, 15(3): 1-10. |
李吉跃. 1989. PV技术在油松侧柏苗木抗旱性研究中的应用. 北京林业大学学报, 11(1): 3-11. |
李良厚, 贾志英, 付祥健. 1999. 土壤水分胁迫下苗木水分参数变化的研究. 河南农业大学学报, 33(1): 92-99. |
刘光崧. 1996. 土壤理化性质分析与剖面描述. 北京: 中国标准出版社, 9-15.
|
刘友良. 1992. 植物水分逆境生理. 北京: 农业出版社, 139-141.
|
毛达如. 1994. 植物营养研究方法. 北京: 北京农业大学出版社, 7-12.
|
阮成江, 李代琼, 姜俊, 等. 2000. 半干旱黄土丘陵区沙棘的水分生理生态及群落特性研究. 西北植物学报, 20(4): 621-627. |
汤章城. 1983. 植物干旱生态生理的研究. 生态学报, 3(3): 196-204. |
王万里. 1984. 压力室在植物水分状况研究中的应用. 植物生理学通讯, (3): 52-57. |
杨敏生, 裴保华, 于冬梅. 1997. 水分胁迫对毛白杨杂种无性系苗木维持膨压和渗透调节能力的影响. 生态学报, 17(4): 364-370. |
张建国, 李吉跃, 姜金璞. 1994. 京西山区人工林水分参数的研究(Ⅰ). 北京林业大学学报, 16(1): 1-11. |
张建国, 李吉跃, 姜金璞. 1994. 京西山区人工林水分参数的研究(Ⅲ). 北京林业大学学报, 16(4): 46-53. |
周以良, 董世林, 聂绍荃. 1986. 黑龙江树木志. 哈尔滨: 黑龙江科学技术出版社.
|
Boucher J F, M unson A D, Bernier P Y. 1995. Foliar absorption of dew influences shoot water potential and root growth in Pinus strobes seedlings. Tree Physiology, (15): 819-823. |
Cheung Y W S, Tyree M T, Dainty J. 1975. Water relation's parameters on single leaves obtained in a pressure bomb and some ecological interpretations. Can J Bot, 53: 1342-1346. DOI:10.1139/b75-162 |
Donald B Z, lee R, Jay H K. 2001. Variation in water relations characteristics of terminal shoots of Port-Orford-cedar(Chamaecyparis lawsoniana)seedlings. Tree Physiology, (21): 743-749. |
Maria J C, Dulce C, Maria M D. 2001. Response to seasonal drought in three cultivars of Ceratonia siliqua:leaf growth and water relations. Tree Physiology, (21): 645-653. |
Grossnickle S C. 1989. Shoot phenology and water relations of Picea glauca. Can J For Res, (19): 1287-1290. |
John E M, Kurt H J. 1999. Shoot water relations of mature black spruce families displaying a genotype ×environment interaction in growth rate Ⅱ.Temporal trends and response to varying soil water conditions. Tree Physiology, (19): 375-382. |
John E M, Kurt H J. 2001. Shoot water relations of mature black spruce families displaying a genotype×environment interaction in growth rate Ⅲ.Diurnal patterns as influenced by vapor pressure deficit and internal water status. Tree Physiology, (21): 579-587. |
Jones M M. 1981. Mechanism of drought resistance.In:Palag L G, Aspinall D.Physiology and Biochemisty Of Drought Resistance in Plants. Sydney :Academic Press: 15-37. |
Jordi M, Joan G. 1997. Effects of water stress cycles on turgor maintenance processes in pear leaves(Pyrus communis). Tree Physiology, (17): 327-333. |
Kozlowski T T.王世绩译.水分供应与树木的生长.林业文摘, 1983, (2): 1~11, (3): 1~5
|
Kurt H J, John E M. 1999. Shoot water relations of mature black spruce families displaying a genotype ×environment interaction in growth rate Ⅰ.Family and site effects over three growing seasons. Tree Physiology, (19): 367-374. |
Schulte P J, Hinckley T M. 1985. A comparison of pressure-volume curve data analysis techniques. J Exp Bot, (36): 1590-1602. |
White D A, Beadle C L, Worledge D. 1996. Leaf water relations of Eucalyptus globulus ssp.globulus and E.nitens:seasonal, drought and species effects. Tree Physiology, (16): 469-476. |
Wilson J R. 1980. Adaption to water stress of the leaf water relation of four tropical species. Aust J Plant Physiol, (7): 208-220. |
Zine E A, Bernier M C, Bernier P Y, et al. 1993. Control of pressure-chamber and rehydration-time effects on pressure-volume determination of water relation parameters. Can J Bot, (71): 1009-1015. |