 2003, Vol. 39
2003, Vol. 39文章信息
- 翟洪波, 李吉跃, 聂力水.
- Zhai Hongbo, Li Jiyue, Nie Lishui.
- 油松的水力结构特征
- THE HYDRAULIC ARCHITECTURE CHARACTER OF PINUS TABULAEFORMIS
- 林业科学, 2003, 39(2): 14-20.
- Scientia Silvae Sinicae, 2003, 39(2): 14-20.
-
文章历史
- 收稿日期:2001-05-10
-
作者相关文章



2. 北京林业大学资源与环境学院 北京 100083
2. College of Resource and Environment, BFU Beijing 100083
水力结构是指植物在特定的环境条件下, 为适应生存竞争的需要所形成的不同形态结构和水分运输供给策略(李吉跃等, 2000)。自Dixon (1914)提出树液上升的内聚力学说(cohesion theory)以后, 随着研究的不断深入, 人们对水分在树体内运动的基本原理有了比较深刻的认识(Huber, 1928; 1956; Van Den Honert, 1948; Scholander, 1972)。然而在国际上, 直到20世纪70年代, 才开始对树木的水力结构(hydraulic architecture)进行研究(Zimmermann, 1971; 1978;Tyree et al., 1975; 1983;1991;Sperry et al., 1987)。以后的20多a来, 对水力结构的研究日趋深入, 如对部分双子叶树木(Zimmermann et al., 1983)和针叶树(Tyree et al., 1983; Ewers et al., 1984; Maurizio et al., 1997; Spicer et al., 1998)的水力结构的研究; Wang等(1992)对北方43个温带树种在冬季水力导度(Kh)的变化的研究等。这些工作从木质部结构、矿质营养(Harvey et al., 1997)、病理(Joseph et al., 1998)、遗传(Vander et al., 1997)、水分动态移动(Melvin, 1988; Thomas, 1997)等不同角度对树木的水力结构进行了有益的探讨, 但目前尚未见到关于相对分枝级、分枝方向、冠层等因素与树木水力结构关系的研究。在国内, 李吉跃等(1988; 1991; 1994; 1995)从20世纪80年代开始对中国北方主要造林树种的耐旱特性及其机理进行了大量研究, 张硕新等(1996)已注意到木质部空穴和栓塞对树木耐旱性的严重影响, 但对树木水力结构的研究尚属空白。
国家自然科学基金课题“我国北方主要造林树种水力结构模型的研究”, 就是针对北方主要造林树种中有代表性的树种来阐明其水力结构特征。这些树种既有裸子植物, 又有被子植物, 而且它们在木质部结构、树冠形态、耐旱机理及耐旱能力等方面都存在着较大的差异, 这对探讨我国造林树种的水力结构特征是有利的。本文仅对油松的水力结构特征进行分析和研究, 旨在深入了解其水分运输的内在规律, 更好地阐明其水分生理特性和耐旱的生态策略, 为抗旱造林树种的选择提供科学的依据。
1 实验材料与方法 1.1 实验材料实验材料油松(Pinus tabulaeformis)来自北京林业大学妙峰山教学实验林场。分别于1999年夏、秋季节(7~9月), 在尚未郁闭的油松林分中选择4株幼树(8~10 a), 在树干上标记分枝方向。清晨日出前, 用锯自地茎处将树木截断, 立即浸入水桶中, 然后迅速运回实验室, 途中注意保水并切勿损伤枝杈。
1.2 实验方法树木运回实验室后, 用标签将全树编号, 详细记录主干和各分枝的长度、侧枝的分枝角度和侧枝的方向(用东E, 南S, 西W, 北N, 东南ES, 西北WN等表示), 汇制成图。在全树中每隔20 cm均匀选取不同部位的枝段, 特别注意选取一定比例分杈基部的枝段(即限速区的枝段), 以便和与其对应的正常部位的枝段(即非限速区的枝段)进行比较。用卡尺测定枝段的长度(用Lz表示)和直径(用d表示), 直径取茎断面上相互垂直的2个方向数值的平均值, 由此推导出的茎段横截面积可以认为是功能木质部的面积, 因为油松是无孔材, 其幼树的心材比例很小或无, 可以忽略不计。同时, 记录所测定茎段水分运输的高度(自地面处至所测定茎段处的垂直距离, 用H表示)和距离(水分自地面处至所测定茎段处所经过的实际长度, 用D表示)。用烘干法测定所测枝段末端的叶干物质质量(用W1表示)。叶面积用公式(1)计算(李家龙, 1985) :
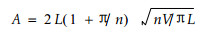
|
(1) |
式中, A为叶面积, V为针叶体积(用排水法测定), n为每束针叶数, L为针叶长度。本文给出叶面积(cm2)和叶干物质质量(g)之间的转换关系:A=165W1。
本文所使用的导度的测量方法比Zimmermann (1978)有所改进, 称之为“改良的冲洗法”。根据Sperry等(1988)的试验结果, 冲洗液使用10 mmol·L-1的草酸溶液, 以免导度在测量过程中下降。压力梯度(△P)定义为水压(MPa)除以枝段长度(Lz), 导水率(Kh)定义为所测茎段单位时间、单位压力梯度下的溶液通过量(gm·MPa-1min-1)。小枝水势在黎明前用压力室测定。
2 结果与分析 2.1 小枝水势的变化图 1表明, 实验期间各样树每天平均小枝水势变化不大, 在-0.1~-0.55 MPa之间。而在干旱条件下, 油松开始发生水分胁迫的临界小枝水势值为(-1.00±0.2) MPa1), 说明尽管季节不同, 但由于自根茎处截断的油松被浸在水中后, 仍能在较长时间内通过木质部吸水供给各部分枝叶利用, 所以各季节供试样树的水分供给状况类似, 均属正常水分条件。这样, 可以将4株不同季节的样树看作是在相同立地条件和小枝水势条件下的4次重复。
1) 李吉跃.太行山区主要造林树种耐旱特性的研究.北京林业大学, 博士论文,1990, 25.

|
图 1 实验期间小枝水势的变化 Fig. 1 Change of twig water potential during experiment |
导水率是树木水力结构中的重要参数之一, 是指茎段在单位压力梯度下、单位时间内的流量, 导水率标志着茎段导水能力的强弱。研究结果表明, 导水率的大小明显受到茎段所在区域的影响。在树体中, 分杈的部位, 即枝迹处, 称之为限速区, 其它部位则称之为非限速区。由图 3可见, 在限速区, 茎段直径(变动在4.3~38.66 mm之间)一般较同一分枝分杈后若干cm后的正常部位(即非限速区, 茎段直径变动在3.76~36.66 mm之间)大, 但限速区的导水率(变动在0.124 1~23.050 3 gm·MPa-1min-1之间)却较相应的非限速区(变动在0.136 6~45.658 6 gm·MPa-1min-1之间)小。在郁闭的林分中, 树木需要和其它相邻的树木竞争光照和空间, 因此, 对喜光树种来说, 顶端的枝条是至关重要的。限速区的存在, 增大了在分枝特别是多级分枝中水分运输的阻力, 从而使油松树体在水分分配上有利于主干和顶梢, 这显然对个体的生存竞争有利。在非限速区, 导水率主要受茎段直径大小的影响, 茎段直径越大, 导水率越高, 用多项式模拟茎段直径与导水率之间的关系, 基本上能得到比较满意的结果, Kh=0.080 1d2-0.527 4d+0.417 9 (n=300, r=0.949 5)。

|
图 3 导水率的变化 Fig. 3 Change of hydraulic conductivity (a)限速区和非限速区对应茎段导水率对比Comparison of corresponding stem segment hydraulic conductivity between constriction and non-constriction areas; (b)非限速区茎段直径和茎段导水率的关系Relations between stem diameter and hydraulic conductivity in non-constriction area (Y=0.080 1x2-0.527 4x+0.417 9 R2=0.901 5).①非限速区Non-constriction area; ②限速区Constriction area. |
相对分枝级是相对于树木枝、干的分枝层次而言的, 树木的主干为0分枝级, 直接着生于主干上的枝条为1分枝级, 直接着生于1分枝级上的枝条为2分枝级, 依次类推(如图 2), 一般来说, 油松幼树的最大相对分枝级不超过3。图 4a~c说明随着相对分枝级的增加(从0~3), 茎段平均直径和茎段平均导水率都逐渐减小(平均直径从34.21至4.29 mm; 平均导水率从98.954 3至0.168 3 gm·MPa-1min-1), 其中以0分枝级(即主干)和1分枝级间的变化最大(平均直径相差24.93 mm, 平均导水率相差96.038 1 gm·MPa-1min-1)。同时, 随着相对分枝级的增加(从1~3), 水分平均运输高度逐渐减小(从148 cm至52 cm), 但水分平均运输距离变化不大(从198 cm至166 cm), 这说明越是下层的枝条, 分枝次数越多, 冠幅越大, 由于3分枝级的枝条大都位于下层树冠, 其水分平均运输距离因树高的降低而减少。

|
图 2 相对分枝级示意图 Fig. 2 Schematic diagram on relative ramification rate |

|
图 4 水力结构各参数随相对分枝级的变化 Fig. 4 Changes of some hydraulic parameters with relative ramification rate (a) The average diameter (d); (b) The average hydraulic conductivity (Kh); (c) The average height (H) and distance (D) of water transportation; (d) The average leaf specific conductivity (LSC); (e) The average specific conductivity (Ks); (f) The average Huber value (HV).下同。The same below. |
|
|
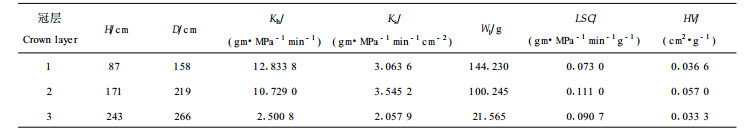
图 4d~f表明随着相对分枝级的增加(从0~3), 茎段平均叶比导率(LSC)、平均比导率(Ks)和平均胡伯尔值(HV)都减小。其中, 主干的平均LSC为0.165 gm·MPa-1min-1g-1, 是3分枝级(0.037 9 gm·MPa-1min-1g-1)的4倍以上; 主干的平均Ks为7.871 4 gm·MPa-1min-1cm-2 , 是3分枝级(1.199 gm·MPa-1min-1cm-2 )的6.6倍, 但HV相差并不是很大, 从主干到3分枝级, 其平均HV分别为0.424, 0.395, 0.352和0.227 cm2·g-1。根据水力结构的基本理论(李吉跃等, 2000), 图 4说明随着枝条相对分枝级的增加, 枝条的平均直径减小, 平均导水率降低, 平均茎段末端叶供水状况越来越差, 茎段本身的输水效率越来越低, 输水阻力不断加大, 供养单位质量的叶所积累的茎段组织则越来越少。这意味着油松幼树在水分胁迫的情况下, 首先受害的将是那些经过多次分枝、较细的末端枝条, 即高分枝级中的枝条, 而尽可能保存主干和力枝等低分枝级的粗壮部分, 油松的这一水力结构特征, 显然有利于其在水分逆境下保护那些已生长多年的光合积累较大的器官。
2.4 冠层对水力结构的影响根据树冠的总长度, 将树冠等分为3份, 1/3冠高以下为第1冠层, 1/3~2/3冠高为第2冠层, 2/3以上为第3冠层, 按冠层求算出各实测值和参数的平均值得表 1。从表中可以看出, 随着冠层的增加, 所测茎段的平均高度和平均运输距离都有所增加, 但茎段末端的平均叶干物质质量减小。比导率(Ks)以第2冠层最高, 平均值为3.545 2 gm·MPa-1min-1cm-2 , 其次是第1冠层, 第3冠层最低, 平均值为2.057 9 gm·MPa-1min-1cm-2 ;这是因为第2冠层枝条分杈次数, 即平均相对分枝级小于第1层, 而输水系统, 即木质部管胞的成熟度优于第3层, 这意味着在全树中, 第2冠层的功能木质部平均输水效率最高。叶比导率以第2冠层最高, 其平均值为0.111 gm·MPa-1min-1g-1, 第1冠层最低, 其平均值为0.073 gm·MPa-1min-1g-1, 第3冠层居中; 说明第2冠层茎段末端叶部的平均供水情况最好, 第1冠层的木质部成熟程度优于第3层, 但其平均相对分枝级最大, 所以茎段末端叶的平均供水状况反不如第3层好。各冠层中, 以第2冠层的胡伯尔值最大, 为0.057 cm2·g-1, 明显高于第1和第3冠层, 但第1和第3冠层的平均胡伯尔值很接近, 分别为0.036 6 cm2·g-1和0.033 3 cm2·g-1, 说明树冠中层供养单位质量叶生存所积累的茎段组织最大。
油松林分中, 下层树冠平均相对分枝级最大, LSC值最小, 叶的光照水分供给条件最差, 这意味着在水分胁迫的情况下, 下层树冠中经多次分枝的末端细小枝条会被首先淘汰。
2.5 分枝方向对水力结构的影响将所测茎段的参数按不同方向归类, 树干方向定为0, 归纳后得图 5。从图中可见, 主干(即0方向)的平均直径、LSC、Ks和HV明显高于其它各个方向, 分别为28.7 mm, 0.116 2 gm·MPa-1min-1g-1, 6.826 4 gm·MPa-1min-1cm-2 , 0.070 3 cm2·g-1; 除主干外, 南向的平均直径、LSC、Ks和HV较高, 分别为7.5 mm, 0.093 8 gm·MPa-1min-1g-1, 2.475 6 gm·MPa-1min-1cm-2 和0.044 9 cm2·g-1; 其次是西南和东南向, 其它各方向则相对较低, 其中以北向最低, 其平均直径、LSC、Ks和HV分别为5.52 mm, 0.040 3 gm·MPa-1min-1g-1, 1.989 7 gm·MPa-1min-1cm-2 和0.024 5 cm2·g-1, 只相当于南向各对应参数的73%, 43%, 36%和55%。与主干比, 则差别更大, 各参数的差别一般在300%以上。

|
图 5 水力结构参数随分枝方向的变化 Fig. 5 Changes of hydraulic architecture parameters with branch directions |
主干是树体的中心, 树木中任何一部分的水分供应都必须首先经过主干才能到达。上述结果说明油松主干不仅是全树中最粗壮的部分, 也是输水效率最高的部分, 在平均蒸腾流密度下, 其输水阻力最小, 对单位叶量的茎组织积累明显高于其它各方向; 其它各方向之间, 阳面枝条(如南向、东南和西南向)的平均直径、LSC、Ks和HV较阴面(如北向、东北和西北向)高。表现在生物学特性上, 油松则具有干性强, 喜光等生物学特性。
3 讨论与结论传统的单位管模型(Shinozaki et al., 1964)把植物木质部看作是均一单位管的集合, 每1个单位管支持1个单位的叶生长。因为人们观察到木质部横截面积一般正比于其末端的叶量, 这一模型被广泛的认可(Smith et al., 1981; Roges et al., 1979)。单位管模型在通过木质部横截面积预测叶面积, 理解树木生长和资源分配等方面确有其实用性(Ewers et al., 1984)。但从水分生理的角度, 本文的实验精确地验证了单位管模型的不正确性。因为在单位管模型中认为树木木质部茎段不论大小位置如何, 都具有相同的给末端叶供水的能力, 即树木各部分的LSC、Ks和HV都相同。但实质上树木不同冠层、不同相对分枝级、不同方向的枝条都存在着差异。
对油松水力结构的研究, 有利于深入地认识其生物学特性和水分供应策略。根据上述研究分析, 得到如下结论:油松树体中, 主干的LSC值最高, 各级分枝中, 随着相对分枝级的增加, 叶比导率逐渐降低, 这使得油松无论是在正常水分条件下, 还是在水分胁迫的情况下, 都能首先保证主干和力枝的水分供给, 这种水分供给策略使油松在逆境下, 能尽可能保留那些光合积累较大的器官。限速区的存在是造成各级主侧枝水分分配差异的原因之一, 油松这样的水分分配特点有利于其个体的生存竞争。油松各冠层中, 下层树冠平均相对分枝级最大, LSC值最小, 叶的光照水分供给条件最差, 这意味着在水分胁迫的情况下, 下层树冠中经多次分枝的末端细小枝条会被首先淘汰。包括主干在内的各方向枝条中, 以主干的水分供给状况最好, 明显优于其它方向的枝条, 而在其它方向中, 阳面枝条校为粗壮, 其水分供给状况好于阴面的枝条, 体现在生物学特性上, 油松具有干性强、喜光的生物学特性。
另外, 在研究相对分枝级、冠层和方向等因素对水力结构的影响时, 本文没有考虑到根系对水力结构的影响, 也未能充分考虑蒸腾强度等环境因子的作用, 这为进一步动态模拟水分在树体中的运输带来不便。要真正弄清树木的水力结构特征, 必须将植物放在SPAC (土壤-植物-大气连续体)中, 结合大气、土壤等环境因子, 综合分析。因此下一步的工作应侧重于对野外活体植物的试验。
李吉跃. 1988. 油松侧柏苗木抗旱特性初探. 北京林业大学学报, 10(2): 23-30. |
李吉跃, 翟洪波. 2000. 木本植物水力结构与抗旱性. 应用生态学报, 11(2): 301-305. DOI:10.3321/j.issn:1001-9332.2000.02.037 |
李吉跃, 张建国, 姜金璞. 1993. 北方主要造林树种耐旱机理及其分类模型的研究(Ⅰ). 北京林业大学学报, 15(3): 1-9. DOI:10.3321/j.issn:1000-1522.1993.03.017 |
李吉跃.太行山主要造林树种耐旱特性的研究(Ⅰ)~(Ⅵ).北京林业大学学报, 1991, (增刊1~2): 1~24;230~280
|
李家龙(摘译).快速测算针叶面积的方法.林业科技通讯, 1985, (10): 封三
|
张建国, 李吉跃, 姜金璞. 1994. 京西山区人工林水分参数的研究(Ⅰ)-(Ⅲ). 北京林业大学学报, 16(1): 1-12. |
张硕新, Richter H. 1996. 浆果紫杉木质部恢复状况的研究. 西北林学院学报, 11(1): 5-8. |
Dixon H H. 1914. Transpiration and the ascent of sap in plants. London: Macmillan.
|
Ewers F W, Zimmermann M H. 1984. The hydraulic architecture of balsam fir(Abies balsamaea). Physiol Plant, 60: 453-458. DOI:10.1111/j.1399-3054.1984.tb04911.x |
Harvey H P, Driessche V D. 1997. Nutrition, xylem cavitation and drought resistance in hybrid popular. Tree Physiology, 17: 647-654. DOI:10.1093/treephys/17.10.647 |
Huber B. 1956. Die gefassleitung.In:Ruhland W.Encyclopedia of Plant Physiology. Springer, Berlin: 541-582. |
Huber B. 1928. Weitere quantitative Untersuchungen ǜber das Wasserleitungssystem der PFLANZEN. Jahrbǜcher fǜr Wissen-schaftlice Botanik, 67: 538-545. |
Joseph G, Kelsey R G, Thies W G. 1998. Hydraulic conductivity in roots of ponderosa pine infected with black-stain(Leptographium wageneri)or annosus(Heterobasidion annosum)root disease. Tree physiology, 18: 333-339. DOI:10.1093/treephys/18.5.333 |
Maurizio M, Grace J, Fioravanti M. 1997. Biomechanical and hydraulic determinants of tree structure in Scots pine:anatomica characteristics. Tree Physiology, 17: 105-113. DOI:10.1093/treephys/17.2.105 |
Melvin J A. 1988. Water Flow in Plants. Longman, London. |
Rogers R, Hinckley T M. 1979. Foliar weight and area related to current sapwood area in oak. Forestry Science, 25: 298-303. |
Scholander P F. 1972. Tensile water. American scientist, 60: 584-590. |
Shinozaki. 1964. A quantitative analysis of plant form-the pipe model theory Ⅰ. Basic analysis.Jap J Ecol, 14(3): 97-105. |
Smith R A, Whitehead D, Jarvis P J. 1981. The contribution of stored water to transpiration in Scots pine. Plant, Cell and Environment, 2: 309-317. |
Sperry J S, Donnelly J R, Tyree M T. 1988. A method for measuring hydraulic conductivity and embolism in xylem. Plant, Cell and Environment, 11: 35-40. DOI:10.1111/j.1365-3040.1988.tb01774.x |
Sperry J S, Holbrook N M, Zimmermann M H, et al. 1987. Spring filling of xylem vessels in wild grapevine. Plant Physiol, 83: 414-417. DOI:10.1104/pp.83.2.414 |
Spicer R, Gartner B L. 1998. Hydraulic properties of Douglas-fir(Pseudotsuga menziesii)branches and branch halves with reference to compression wood. Tree Physiology, 18: 777-784. DOI:10.1093/treephys/18.11.777 |
Thomas F. 1997. Simulation of water flow in the branched tree architecture. Silva Fennica, 31(3): 275-284. |
Tyree M T, Caldwell C, Dainty J. 1975. The water relations of hemlock(Tsuga canadensis)Ⅴ.The localization of resistances to bulk water flow. Canadian Journal of Botany, 61: 2015-2111. |
Tyree M T, Ewers F W. 1991. The hydraulic architecture of trees and other woody plants. New Phytol, 119: 345-360. DOI:10.1111/j.1469-8137.1991.tb00035.x |
Tyree M T, Graham M E, Cooper K E, et al. 1983. The hydraulic architecture of Thuja occidentalis. Can J Bot, 61: 2105-2111. DOI:10.1139/b83-226 |
Van Den-Honert T H. 1948. Water transport in plants as a catenary process. Discussions of the Faraday Society, 3: 146-153. DOI:10.1039/df9480300146 |
Vander G, Van Boxel. 1997. Water and surface energy balance model with a multilayer canopy representation for remote sensing purposes. Water Resour Res, 25(5): 949-971. |
Wang J, Ives N E, Lechowicz M J. 1992. The relation of foliar phenology to xylem embolism in trees. Functional Ecology, (6): 469-475. |
Zimmermann M H, Brown C L. 1971. Trees:Structure and Function. Springer-Verlag, Berlin, New York. |
Zimmermann M H, Sperry J S. 1983. Anatomy of the palm Rhapis excelsa Ⅸ.Xylem structure of the leaf insertion.. J Amold Arbor Harv Univ, 64: 599-609. |
Zimmermann M H. 1984. Functional xylem anatomy of angiosperm trees.In:Baas P(ed).New Perspective in wood Anatomy. M artinis Nijhoff Publishers, The Hague, Boston: 59-70. |
Zimmermann M H. 1978. Hydraulic architecture of some diffuse-porous trees. Canadian Journal of Botany, 56: 2286-2295. DOI:10.1139/b78-274 |