 2003, Vol. 39
2003, Vol. 39文章信息
- 吴大荣, 朱政德.
- Wu Darong, Zhu Zhengde.
- 福建省罗卜岩自然保护区闽楠种群结构和空间分布格局初步研究
- PRELIMINARY STUDY ON STRUCTURE AND SPATIAL DISTRIBUTION PATTERN OF PHOEBE BOURNEI IN LUO BOYAN NATURE RESERVE IN FUJIAN PROVINCE
- 林业科学, 2003, 39(1): 23-30.
- Scientia Silvae Sinicae, 2003, 39(1): 23-30.
-
文章历史
- 收稿日期:1999-03-16
-
作者相关文章


2. 南京林业大学 南京 210037
2. Nanjing Forestry University Nanjing 210037
闽楠〔Phoebe bournei(Hemsl.)Yang〕为我国珍稀濒危树种,其树干通直、木材纹理结构细密,削面光滑美观,有香气,为高级建筑、家具、雕刻和精密木模的珍贵用材树种。由于人为砍伐、自身生物学特性以及自然环境等综合因素作用,致使闽楠资源接近枯竭,现仅在浙南、闽、赣、鄂西、湘、粤北、桂和黔东等海拔200~1 000 m的山地常绿阔叶林中有零星分布。因此,开展闽楠种群生态学研究对认识和保护这一珍稀树种具有重要意义。福建省闽楠林保存较完整的仅见于沙县、南平和政和。1983年福建省三明市沙县罗卜岩建立了闽楠种质资源保护区。1994年我们曾对保护区内闽楠林进行种群调查测定,1997年进行补充调查。本文为1997年调查测定结果。
1 自然环境和植被概况福建省三明市沙县罗卜岩自然保护区位于东经117°34′30″,北纬26°26′30″,主峰海拔642 m,相对高度285 m。气候温暖湿润,属典型的中亚热带气候,年均温19.5℃,最冷月1月份均温8.5℃,最低温-7.1℃,最热月7月份均温28.4℃,最高温40.1℃。年降水量1 643.2 mm,无霜期297 d。基岩多为花岗岩,土壤为红壤和红黄壤,土层厚度多在80 c m以上,较陡的山坡基岩常裸露。土表枯落物层较厚。区内亚热带常绿阔叶林以闽楠林、栲类林为主;亚热带常绿、落叶阔叶混交林有喜树(Camptotheca acuminata)-鹿角栲(Castanopsis lamontii)林,光皮桦(Betula luminifera)-台湾冬青(Ilex formosana)-闽楠林等;此外,还有竹、马尾松(Pinus massoniana)林等(邹惠渝等,1997)。
2 研究方法 2.1 样地环境情况调查在全面勘察的基础上,选择哨所(1)、旗坑下(2)、旗坑上(3)、涵洞顶(4)和坝头(5)等5块样地,样地面积各为1 600 m2,记录每样地群落类型、海拔高度、坡向、坡度、坡位、土壤厚度、土壤pH值、土壤含水量、土壤有机质含量以及群落透光度(表 1)。用5 m×5 m相邻格子法(陆阳,1989)进行每木调查,记录所有闽楠个体。
|
|

闽楠材质坚硬,用生长锥钻取木芯十分困难,外部特征也难精确测定生长年龄。本文用立木级结构代替年龄结构分析种群动态。大小结构按两种方式处理(刘智慧,1990):胸径2.5 cm以下个体按树高分为3级:Ⅰ级高1~33 cm,Ⅱ级高33~100 cm,Ⅲ级高大于100 cm;胸径大于2.5 cm的个体,按胸径大小分级,每增加5 cm为1级(图 1),并绘制闽楠种群存活曲线(如图 1-f)。

|
图 1 闽楠种群大小结构与存活曲线 Fig. 1 Size structures and survival curves of Phoebe bournei population ○—○样地1 Plot 1;△—△样地2 Plot 2;·—·样地3 Plot 3;×—×样地4 Plot 4;——样地5 Plot 5. |
对地径大于2.0 cm的闽楠个体空间分布格局进行拟合,离散分布理论拟合采用泊松分布、负二项分布和奈曼分布等方法(Cox, 1979;Pielou, 1969);聚集强度指标有负二项式指数K、聚块性指挤m*/m 、丛生指标I和平均挤度m*等(皮洛,1988;赵志模等,1984)(表 2)。群落类型有常绿阔叶林群落和常绿落叶阔叶混交林群落(图 3)。
|
|

|
|


|
图 3 不同群落类型闽楠种群空间分布格局负二项指数K Fig. 3 Negative binomial distribution index K in different communities |
把5块样地中闽楠种群个体按曲仲湘(1952)标准划分如下:
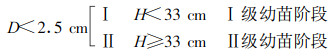
|
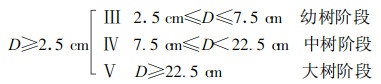
|
用方差/均值比率法(皮洛,1988;赵志模等,1984)对不同发育阶段的闽楠种群进行空间分布格局类型分析,并计算其聚集强度指标(表 3)。
2.3.3 种群空间分布格局分析采用Greig-smith 方法(1952)和Kersaw 方法(1970)对闽楠种群空间分布格局进行分析(表 4)。
3 结果与分析 3.1 闽楠种群结构、静态生命表和存活曲线将5个样地闽楠种群的数量统计分级如图 1。从图 1的a~e可见,种群结构都属于基部宽而顶部狭窄的金字塔型,表现为有丰富的幼苗贮备。闽楠种群幼苗贮备(包括Ⅰ,Ⅱ和Ⅲ级幼苗)所占比例分别为77.588%、99.348%、83.226%、93.514%和90.353%。图 1-f为闽楠种群存活曲线。从图 1-f看,闽楠种群存活曲线的共同特征表现为:从Ⅰ级幼苗到Ⅱ级幼苗发育过程中,出现了死亡高峰期。样地1~5幼苗死亡率分别为45.0%、99.2%、78.6%、94.5%和93.9%。在不同林型中,存活曲线表现出较大差异。常绿阔叶林两样地(样地4、5)中闽楠种群进入稳定阶段的径级较为一致,大约为30 cm。其它3个常绿落叶阔叶林样地的闽楠种群存活曲线差异较大。样地2中闽楠种群已进入稳定阶段。样地1和3的闽楠种群尚未呈现稳定趋势,其原因是闽楠尚未挤入林冠上层,占主导地位,林冠上层仍由落叶树种如光皮桦(样地3)和喜树(样地1)等占绝对优势,闽楠种群仍处于激烈竞争状态,数量还在变动,呈下降趋势。
在不同演替群落中,闽楠种群大小结构呈现出规律变化。图 2为不同演替群落中闽楠种群大小结构中最大径级随乔木层常绿树种重要值/落叶树种重要值变化情况。从图 2可见,随着常绿树种重要值的增加,闽楠种群最大径级也增大。这是由于在落叶树占绝对优势的群落(如样地1)中,闽楠种群侵入时间不长,表现为最大径级≤17.5 cm。之后,当闽楠种群逐渐扩大,达到即将与落叶阔叶树种平分主林层时(如样地3),闽楠种群已经历了较长时期,最大径级达42.5 cm。根据解析木资料(邹惠渝等,1997),闽楠成熟林年龄≥170 a,相应的胸径约42.40 cm。可见,样地1、3闽楠种群增长处于旺盛期,未达成熟型,未有成熟种群的结构特征,即尚缺乏42.50 cm以上的径级。随着群落的演替,当闽楠等常绿阔叶树种在林冠层占较大成势时(如样地2),种群经历了更长时期,也有较大的径级,如最大径级达47.5 cm。当闽楠等常绿阔叶树种在林冠层中占绝对优势时(如样地4和5),最大径级达52.5 cm,在样地2、4、5中,闽楠种群径级已超过42.5 cm,种群进入了成熟型阶段。但从5个样地情况看,闽楠种群都未进入衰退型阶段,表明罗卜岩自然保护区闽楠种群依然呈稳定增长型或进入成熟型阶段。综上所述,随着群落演替的进行,闽楠种群的大小结构呈现动态变化。

|
图 2 5个样地乔木重要值比(常绿/落叶)和闽楠种群最大径级 Fig. 2 Ratio of importance value (evergreen/deciduous) and maximum diameter class of Phoebe bournei population in five plots ○—○ 重要值比 Ratio of importance value;·—· 最大径级 Maximum diameter class. |
从表 3可见,5块样地闽楠种群空间分布格局以负二项分布为主。彭少麟、王伯荪(1984)认为负二项式的参数K值,能作为衡量集群程度的指标,对研究植物种的集群分布较为有效。图 3显示了两种不同群落类型中闽楠种群空间分布格局聚集强度K值的情况。样地1、2、3为常绿落叶混交林群落,样地4和5为常绿阔叶林群落。从图 3可见,常绿阔叶林群落中闽楠种群空间分布聚集强度比其在常绿落叶阔叶混交林中聚集强度低。森林演替过程中种群分布格局产生变化,是由于不同演替阶段的群落内环境不同,随着群落的演替也渐渐改变了周围的环境1),在这过程中,种群的变化反映在种群结构的物种星座图变了(王伯荪等,1983),群落的物种多样性也变了(彭少麟等,1983)。在常绿落叶阔叶混交林中,落叶树种一年一度的落叶形成林窗,林内光照水平较高,种内、种间对光照水平的竞争较为缓和,种群自然稀疏过程较缓慢,空间分布格局聚集度因而能保留较高状态。而在常绿阔叶林中,由于林内光照水平低(表 1),种内、种间对光需求的竞争激烈,种群个体死亡率增加,空间格局也产生变化,由聚集度高向聚集度低转变。
1) 彭少麟,王伯荪.鼎湖山森林群落分析Ⅲ.种群分布格局.见:中国科学院鼎湖山森林生态系统定位研究站编,热带亚热带森林生态系统研究,第1版,1984(2):24~37.
3.2.2 不同发育阶段闽楠种群空间分布格局同一种群在不同径级阶段分布格局有差异。从表 3可看出,闽楠种群分布格局立木级的变化是十分明显的,且显示出较为一致的规律,即:第Ⅰ、Ⅱ、Ⅲ级立木多为集群分布,第Ⅳ、Ⅴ级为随机分布,仅有个别例外。据研究,中生性常绿阔叶树种种群分布格局大多是趋于集群分布,是外界生态因素综合影响的结果,也是种群本身的特性所决定,尤其是与种子的成群散布有关1)。在中亚热带闽楠群落中,每年秋季闽楠种子成熟,在重力作用下密集于母树周围,翌年4~6月幼苗萌发,幼苗仍呈密集状态,这种状态持续到小树阶段。随着种群继续发育,种内和种间竞争加剧,种群个体死亡率提高,种群密度下降,到了中树和大树阶段,种群分布格局发生了明显变化,由集群分布向随机分布转变。种群在幼年阶段集群强度高有利于存活和发挥群体效应,而成年时,由于个体增大,集聚强度降低则有利于获得足够的环境资源(蔡飞等,1997)。
3.2.3 不同群落生境闽楠种群空间分布格局不同群落生境变化对种群分布格局有影响(王伯荪等,1983)。同一种群在不同群落不同海拔高度的分布格局也有差异,陆阳从种群分布格局与海拔高度关系上,发现尽管黄果厚桂壳(Cryptocarya concinna)种群分布格局受海拔梯度明显影响,但多数种群对海拔高度没有表现出分布格局的明显不同(陆阳,1982)。对闽楠而言,不同的海拔高度对闽楠种群空间分布格局也产生一定影响,如低海拔为泊松分布,较高海拔为集群或奈曼分布,但变化不很一致。将表 1和表 2对照发现,在其它生态因子中,对闽楠种群空间分布格局产生明显影响的有坡度和群落透光度。坡度可分3个等级,即群落1,群落2和3,群落4和5,从空间分布格局T值和K值可以明显看出,随着坡度的增加,空间分布格局趋于泊松分布,聚集强度K值减弱,其原因推测为以重力传播为主的闽楠种子落在较平坦的林地后,非动物因子的后传播(postdispers al)(Horvitz et al.,1994)机会减少,从而能较稳定地留在落地时的位置,而较陡的山地,后传播可能性大,破坏了原来可能的分布格局。虽然种子的初始分布格局对其萌发的幼苗的分布并不一定存在密切相关(Houle, 1992),但由群落本身产生的种子分布,由于重力作用则较多地趋于集聚分布1),也影响幼苗的分布格局。从表 1和表 2看,坡位并没有使处于下坡的闽楠种群明显地趋于集群分布。笔者认为,坡位的影响往往与处于样地上方的母树有否,数量多少,分布情况等有关。群落透光度的影响,可能通过林内光照水平影响种群自然稀疏过程。据研究,即使是顶极树种(climax species)在遇到较好光照条件下也能表现出“释放”(release)生长(Thomas, 1989)。闽楠为顶极树种,林内如有较好光照水平,种群自然稀疏作用相应就减缓,空间分布格局仍保留原有集群分布的态势。相反,光水平较低的林内,种群自然稀疏作用较强,使一些个体死亡,分布格局可能因此受影响,从强聚集向弱聚集变化的可能性提高了。光照水平的差异是由群落演替结果产生的,群落演替引起了光照水平差异,光照水平差异引起空间分布格局变化。
3.2.4 闽楠种群空间分布格局分析对小规模的空间异值性即植物种群的分布格局进行分析,可定量描述种群分布格局的强度、规模和纹理(陆阳,1987),而分布格局规模的确认可通过面积大小来指示,这一观点已被广为接受(陆阳,1987;杨持,1982;Greig-Smith, 1979;Kershaw, 1964;1970),但某规模上确认分布格局强度大小却有争议(陆阳,1987)。本文将各样地划分区组为1,2,4,8,16和32等6个,采用Greig-Smith方法和Kersaw方法,对闽楠种群空间分布格局分析如表 4。
|
|
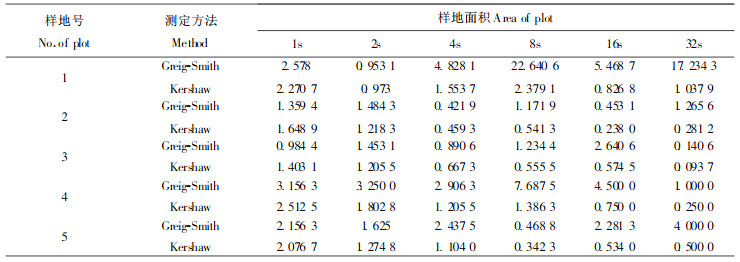
从表 4看出,随着组的大小增加,均方也增加,直至组的大小等于聚块的面积为止。对空间分布格局的分析,Greig-Smith方法在一定程度上消除了野外样方大小造成的影响,并有效地分离分布格局的规模和强度,但该方法也存在区组间隔过大、分析结果依赖初始点和基本样方大小等弱点(陆阳,1987;皮洛,1988;Errington,1973;Ripley,1978;Usher, 1975)。如果要进一步地进行详细分析,有学者建议可用Hill方法(1973)结合应用(陆阳,1987)。Kershaw方法把一强度的标准化度量用以直接指示强度和规模关系,很明显,它没有合适的统计检验,同时,峰值的直观显著性也降低。
3.3 闽楠濒危原因和保护措施初探关于闽楠濒危机制问题,有些学者认为天然更新不良是主要原因(湖南森林编委会,1991),但也有的认为闽楠天然更新良好(浙江森林编委会,1993)。分析以往的资料不难发现,认为闽楠天然更新不良或良好的结论均是文字描述性质的,未见有详细的种群调查数据。通过对罗卜岩闽楠种群详细调查研究,笔者认为从闽楠种群大小结构看,闽楠种群有丰富的幼苗贮备,种群呈稳定增长型或成熟型,并具有巨大的种子库(吴大荣,1997),说明罗卜岩自然保护区现存生境适合闽楠种群繁衍。笔者认为,闽楠濒危的重要原因之一是生态因素,即人为破坏致使种群数量减少。
如何有效地保护珍稀树种闽楠是一个艰巨的任务,目前建立保护区以就地保护方式将有助于闽楠进行天然更新,是保护种质资源的重要手段。例如,福建省南平宝珠大垄的闽楠林,虽然面积不大,自清乾隆33年繁衍至今,历来有保护风尚,相沿成习,至今保存完整而茂盛(福建森林编委会,1993),林下闽楠的幼苗幼树也十分丰富。但由于在保护区和风水林之外,闽楠资源仍然面临人为的威胁,种群数量日趋减少,至使闽楠在分布区已呈零星状(傅立国,1989),生境严重破坏,加之,由于现有保护区大多以孤岛状存在,造成种群分割,种群近亲交配率提高,从而可能降低繁殖力,使种群进一步衰弱,复壮(recovery)的可能性减小(Parcker,1993)。因此,要有效地增加地理种群间个体流和扩大有效种群数目达到长期保护目的,在保护区之间宜建立“绿色走廊”以促进基因交流(郑向忠等,1997),从而达到有效保护,使该濒危树种得以繁衍。在保护的同时还应积极开展引种研究工作,大量繁殖人工种群。
4 结论罗卜岩自然保护区闽楠种群结构呈金字塔型,幼苗贮备丰富;不同群落演替类型中,种群大小结构不同,但随着群落的演替,呈现一定的规律变化;种群存活曲线在Ⅰ级~Ⅱ级苗时期陡然下降,为死亡高峰期;罗卜岩闽楠种群呈稳定增长或正处于成熟阶段,尚未进入衰退阶段。不同群落演替类型中,闽楠种群空间分布格局不同,在常绿阔叶林群落中其空间分布聚集强度比在常绿落叶阔叶混交林群落中低;不同发育阶段空间分布格局存在较大差异,总体趋势由集群向随机分布转变,由聚集度高向聚集度低转变;不同群落生境对种群空间分布格局产生影响,其中群落透光度和坡向影响较为明显。同时,不同区组大小也对空间分布格局产生影响。闽楠种群濒危的重要原因之一是人为砍伐过度,致使其生境破坏,种群数量减少。因此,就地保护和增加种群间基因交流是其重要保护措施。
蔡飞, 宋永昌. 1997. 武夷山木荷种群结构和动态的研究. 植物生态学报, 21(2): 138-148. DOI:10.3321/j.issn:1005-264X.1997.02.005 |
《福建森林》编委会编著.福建森林.第1版.北京: 中国林业出版社, 1993: 23~29, 143~148
|
傅立国主编.中国珍稀濒危植物.第1版.上海: 上海教育出版社, 1989: 177
|
《湖南森林》编委会编著.湖南森林.第1版.长沙: 湖南科学技术出版社, 北京: 中国林业出版社, 1991: 236~238
|
《江西森林》编委会主编.江西森林.第1版.南昌: 江西科学技术出版社, 北京: 中国林业出版社, 1986: 51~66
|
刘智慧. 1990. 四川省缙云山栲树种群结构和动态的初步研究. 植物生态学与地植物学学报, 14(2): 120-128. |
陆阳. 1982. 鼎湖山森林群落数量分析. 生态科学, 1: 74-80. |
陆阳. 1987. 鼎湖山森林植物种群分布格局分析与联结分析. 武汉植物学研究, 5(4): 360-371. |
陆阳. 1989. 南亚热带森林种群分布格局取样技术研究. 植物生态学与地植物学学报, 10(4): 273-282. |
皮洛, E.C.(卢泽愚译).数学生态学.第2版.北京: 科学出版社, 1988: 119~193
|
彭少麟. 1984. 森林群落植物种群分布格局变因探讨. 生态科学: 10-15. |
彭少麟, 王伯荪. 1983. 鼎湖山森林群落分析(Ⅰ).物种多样性. 生态科学, 1: 11-17. |
王伯荪, 彭少麟. 1983. 鼎湖山森林群落分析(Ⅱ).物种联结性. 中山大学学报(自然科学版), 4: 27-35. |
吴大荣. 1997. 福建省罗卜岩自然保护区闽楠种群种子雨研究. 南京林业大学学报, (1): 56-60. |
杨持. 1982. 羊草草原群落水平格局的研究(1)邻接格子样方的应用. 内蒙古大学学报, 2: 245-254. |
邹惠渝, 吴大荣主编.闽楠种群生态学.第1版.北京: 中国林业出版社, 1997: 8~20
|
赵志模, 周新远. 1984. 生态学引论.第1版. 重庆: 科学技术文献出版社重庆分社, 93-125.
|
《浙江森林》编委会编著.浙江森林.第1版.北京: 中国林业出版社, 1993: 182
|
郑向忠, 徐宏华, 陆厚基. 1997. 动物种群遗传异质性研究进展. 生物多样性, 5(3): 210-216. DOI:10.3321/j.issn:1005-0094.1997.03.010 |
Cox G W.(蒋有绪译).普通生态学实验手册.第1版.北京: 科学出版社, 1979: 72~85
|
Errington J C. 1973. The effect of regular and random distributions on the analysis of pattern. J Ecol, 61: 99-105. DOI:10.2307/2258920 |
Greig-Smith P. 1952. The use of random and contiguous quadrats in the structure of plant communities. Ann.Bot, 16: 293-316. DOI:10.1093/oxfordjournals.aob.a083317 |
Greigh-Smith P. 1979. Pattern in vegtation:presidential address to the British Ecology Society. J Ecol, 67: 755-779. DOI:10.2307/2259213 |
Hill M D. 1973. The intensity of spatial pattern in plant communities. J Ecol, 61: 225-235. DOI:10.2307/2258930 |
Houle G. 1992. Spatial relationship between seed and seedling abundance and mortality in a deciduous forest of north-eastern North America. J Ecol, 80: 99-108. DOI:10.2307/2261066 |
Horvitz C C, Schemske D W. 1994. Effects of dispersers, gaps, and predators on dormancy and seedling emergence in a tropical herb. Ecology, 75(7): 1949-1958. DOI:10.2307/1941599 |
Kershaw K A.Quantitative and dynamics Ecology.lst published.London: Edward Arnold ltd., 1964
|
Kershaw K A. 1970. An empirical approach to the estimation of pattern intensity from density and cover data. Ecology, 51: 729-734. DOI:10.2307/1934056 |
Parcker P G, Whiteman H H. 1993. Genetic diversity in fragmented populations of Clemmys Guttata and Chrysmys picta marginata as shown by DNA finger printing. Copeia, 3: 841-846. |
Pielou E C.An Introduction to Mathematical Ecology.New York: Wiley-Intersicence, 1969
|
Ripley B D. 1978. Spectral analysis and the analysis of pattern in plant communities. J Ecol, 66: 965-981. DOI:10.2307/2259308 |
Thomas T Veblen. 1989. Tree regeneration responses to gaps along a transandean gradient. Ecology, (3): 541-543. |
Usher M B. 1975. Analysis of pattern in real and artificial plant populations. J Ecol, 63: 569-585. DOI:10.2307/2258736 |