 2003, Vol. 39
2003, Vol. 39文章信息
- 贺康宁, 张光灿, 田阳, 史常青, 宋吉红.
- He Kangning, Zhang Guangcan, Tian Yang, Shi Changqing, Song Jihong.
- 黄土半干旱区集水造林条件下林木生长适宜的土壤水分环境
- THE SUITABLE SOIL MOISTURE CONDITION FOR FOREST GROWTH IN CATCHMENT AFFORESTATION IN SEMI-ARID REGION ON LOESS PLATEAUS
- 林业科学, 2003, 39(1): 10-16.
- Scientia Silvae Sinicae, 2003, 39(1): 10-16.
-
文章历史
- 收稿日期:2001-04-17
-
作者相关文章


2. 山东农业大学林学院 泰安 271018
2. Forestry College, Shandong Agricultural University Tai'an 271018
对集水整地为基础的径流林业而言,林木生长所适宜的土壤水分条件是估算林木需水量和确定林木水分环境容量的依据和关键。而适宜的水分条件应该是多少,维持林木正常生长,至少获得中度以上的生产量,所需要的水分上、下限有多大,这是节水农林业生产上田间水分管理问题的标准和基础,这个标准需要通过对地区环境和植物种类开展系统的定量研究,以期建立植物节水高产的生产模式。从理论上讲,在大田环境条件下研究土壤水分与植物需水量、水分生产效率及生物量(或是经济产量)之间的关系,是确定植物适生土壤水分条件的最佳途径。尽管目前很难完全控制研究林地的土壤水分环境条件,但通过不同集水技术所形成的不同林地的土壤水分差异,辅以盆栽试验条件下的不同水分梯度观测结果,就可以较准确的探求实生大田林木的基本水分生理生态特性。
1 试验条件和方法 1.1 试验地概况试验地位于山西省吕梁山西麓的方山县峪口镇土桥沟流域(北纬37°36′58″,东经110°0 2′55″)的北京林业大学径流林业试验场。流域内最高海拔1 446 m,试区平均海拔1 200 m左右。该地区属暖温带大陆性季风气候,干燥度1.3。冬春寒冷干燥,秋季凉爽少雨,夏季降雨集中,无霜期140 d。据方山县气象站1975~1992年资料统计,该地区多年平均降水量416 mm,且年内分配非常不均,6~9月份降水占全年的70%以上;多年平均水面蒸发量高达1 857.7 mm,最大蒸发出现在4~6月,表现出典型的北方严重春旱的特征。试验区属典型黄土丘陵沟壑地貌,地表大部分为新生代第四纪马兰黄土所覆盖,土壤为黄绵土,由黄土母质直接发育形成,层次过渡不明显;土层深厚,质地均匀,为中壤土。试验地平均土壤容重1.20 g·cm-3,田间持水量21.0%左右。
1.2 试验方法试验材料选取了本试验场9 a生刺槐(Robinia pseudoacacia)10株,其分别处于3种集水处理、5种密度的5个试验小区(每小区有2株重复),由集水措施和林分密度不同所自然形成的林地土壤水分差异为试验条件。同理,选取了2种集水处理,5种密度的12 a生侧柏(Platycladus orientalis)10株。选用10株施用了保水剂的9 a生(旱园)金冠苹果(Malus pumila),由人为给水产生水分梯度,苹果生理观测在人工给水24~36 h后,当土壤剖面土壤水分渗透稳定后进行。
另外布设了3 a生刺槐和4 a生侧柏苗木(各20株)的盆栽实验,通过人为控制产生水分亏缺梯度,作为低限土壤水分胁迫条件的辅助观测,以验证试验小区在低土壤水分条件观测时的数据结果。
林木蒸腾和光合速率采用生长季典型天气全天观测的方法,每2 h观测一次。蒸腾速率的观测使用LI-1600稳态气孔计,光合作用观测使用LI-6200光合作用测定仪。观测时,在刺槐、侧柏林木树冠南、北方向的中部和上部,在苹果树冠南、北、东、西方向的中、上部,分别确定标准枝,选取健康成叶进行测定;蒸腾速率和光合作用的观测同步进行。对刺槐、侧柏林冠上部叶片蒸腾作用的观测采用剪枝取样法进行(郭柯等,1996)。
对刺槐蒸腾作用的观测,叶片正、反面均进行测定,叶片总蒸腾速率为正、反面之和。叶片总气孔阻力(RSt)计算公式为(康绍忠等,1991):RSt-1= RSa-1+RSb-1。式中:RSt为叶片总气孔阻力,RSa为叶片正面气孔阻力,RSb为叶片反面气孔阻力。
一般情况下,气孔计测定的蒸腾速率高于自然蒸腾值(刘奉觉等,1992;1997),而且水分环境条件越好,误差越大。其与仪器中送入的干空气、仪器内较高的气流流速及较低的界面层阻力有关。因此,本研究由快速称重法(感量为千分之一的天平)与同期相同土壤水分条件下LI-1600稳态气孔计的观测值绘制了校正曲线,对仪器实测值进行了适度校正(刘奉觉等,1988;1992)。
参加土壤水分与林木生理参数关系分析的数据,选自5~9月观测日、晴天9:00~11:00、光量子通量密度为800~1 100 μmol·m-2s-1范围内的观测结果。
林地土壤含水量的观测,采用国产LNW-50A型中子土壤水分仪,在林木生理观测当日同步进行,深度为每间隔20 cm一层。盆栽土壤含水量在采样后由烘干法测定。
有关线性和非线性回归统计计算,采用了目前常用的SPSS统计分析软件。
2 结果与分析就林木生理指标与土壤含水量的关系(重量含水量, %),本研究主要选择了如下内容:
净光合速率(Pn:μmol·m-2s-1)与土壤含水量;蒸腾速率(Tr : mmol·m-2s-1)与土壤含水量;叶片水分利用效率(WUEL:μmol·CO2·mmol H2O-1)与土壤含水量;羧化效率(CE: mol·m-2s-1)与土壤含水量;气孔导度(Cs: cm·s-1)或气孔阻力(Rs: s·cm-1)与土壤含水量;光合速率与叶片气孔导度;蒸腾速率与叶片气孔阻力。
叶片水分利用效率(WUEL)指单位水量通过叶片蒸腾散失时光合作用所同化的CO2量,为光合速率与蒸腾速率的比值(Pn/Tr),是水分利用效率的理论值(刘昌明等,1999);羧化效率采用光合速率与胞间CO2浓度的比值(Pn/Ci)来衡量,其大小表明叶肉因素对Pn的影响程度(邹琪,1994);气孔状况指标分别采用气孔阻力(一定温度下单位水蒸汽量通过单位叶面积由叶肉扩散到叶外空气中所需要的时间)和气孔导度(气孔阻力的倒数表示),气孔导度与CO2或水汽扩散通量呈线性关系,使用起来比较方便,而气孔阻力与扩散通量呈双曲线关系,数据处理不方便。大量研究表明,蒸腾作用及气孔导度都随水分胁迫的发展而降低,是2个反映土壤水分亏缺的最敏感指标。
通过2000年5、7、9月的集中实地观测,得到刺槐、侧柏和苹果生理指标与土壤水分的关系,经统计回归,建立了其数学拟和模型(见表 1)。
|
|
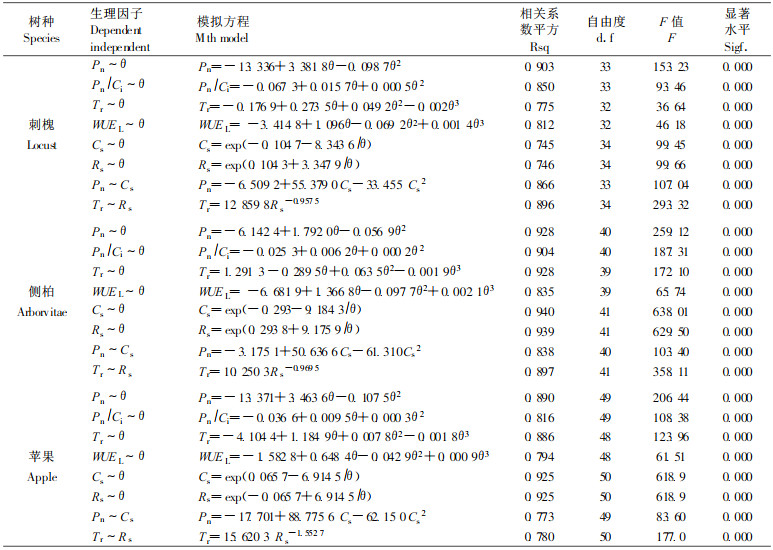

|
图 1 刺槐、侧柏和苹果净光合速率、羧化效率及气孔导度与土壤水分的相互关系 Fig. 1 The relationships between net photosynthesis rate, carboxylation efficiency, stomatic conductivity of Locust, Arborvitae & Apple with soil water content |
已有研究表明,侧柏与刺槐的土壤水分补偿点分别为-5.37 MPa和-3.91 MPa左右(李吉跃,1991);黄土高原主要树种凋萎湿度的研究结果中,刺槐为4.52%,侧柏为3.9%,苹果为4.28%~4.38% (王斌瑞,1988),与本研究的结果基本一致。
从以上分析并参照图 1结果,可以确定:
刺槐光合速率最大值相对应的土壤含水量相当于田间持水量的81.57%,此时的气孔导度Cs在随土壤水分变化的明显转折点0.55 cm· s-1附近;如光合速率维持最高水平的70%以上,其相对应的土壤含水量最低应在10.25%(相当于48.8%的田间持水量)以上,此时的气孔导度Cs恰好在由急变缓的0.4 cm·s-1附近(图 1D)。
侧柏维持最大净光合速率的土壤含水量大约在田间持水量的75.7%左右,比刺槐低6%,此时的Cs在0.41 cm·s-1处;维持70%最大光合速率的土壤含水量应在9.27%以上(相当于44%的田间持水量)。
苹果最适光合作用相对应的土壤含水量相当于田间持水量76.71%,比刺槐低5%,此时的气孔导度Cs在0.7 cm·s-1附近;维持最大光合速率70%以上水平相对应的最低土壤含水量为9.75%,相对应的Cs值位于随土壤含水量递增由急变缓的0.53 cm·s-1处。
2.2 蒸腾作用与土壤水分的关系集水措施改善了林木根系的吸水条件,从而影响到水分由土壤经林木树体输送到叶片的速率,使叶片组织受水分胁迫的概率减小,这一过程的效果可以从叶片的气孔阻力表现出来。图 2表明,由于土壤水分条件对刺槐气孔阻力的影响而最终影响到蒸腾速率,表现出土壤含水量越高,叶片在一天中整体的气孔阻力越低;蒸腾速率随气孔阻力降低而递增。但同一时间的土壤水分对叶片气孔阻力的影响不及太阳辐射强度因子。图 2C为正午与早晚的Tr-Rs曲线,每条拟和曲线的数据中包含4种土壤水分条件下测定的结果,说明太阳辐射强度对Tr-Rs关系的影响程度大于土壤水分。这是因为气孔阻力的大小除受制于叶组织结构外,主要受气候因子的调节而不是土壤水分的供应能力(康绍忠等, 1994)。

|
图 2 不同土壤水分条件下刺槐的蒸腾速率和气孔导度(1998-06-28) Fig. 2 The Transpiration rate and stomatic conductivity of Locust in different SWC |
图 3表明,蒸腾速率与太阳辐射强度同步变化,在林冠的不同部位由于透射的光照强度发生变化,气孔阻力也相应改变,气孔阻力从下到上随光照强度增加而递增;在相同的土壤水分条件下,蒸腾速率随着气孔阻力的降低而增加(图 4)。水分蒸腾在植物体内产生的水势梯度控制着水分吸收和液流上升的速度,在炎热晴朗的天气条件下,蒸腾作用几乎每天都会引起叶片的暂时水分亏缺,特别是当土壤干燥、土壤水势降低使根系吸水滞后于蒸腾失水时,就会发展成持久性的水分亏缺并导致植物萎蔫。实际上,在干旱半干旱地区,由于蒸腾引起的水分亏缺使植物生长受到伤害或引起枯死的现象要比任何其它原因造成的多(王斌瑞等,1996)。若蒸腾作用被减少而又不影响光合作用,则植物的水分利用效率就会显著提高,由干旱造成的伤害也会大为减少。

|
图 3 刺槐林冠层不同光照条件下的蒸腾速率、气孔导度日过程(1998-06-25) Fig. 3 The daily course of transpiration rate and stomatic resistance of locust Canopy in different illuminance |

|
图 4 刺槐、侧柏和苹果蒸腾速率、气孔阻力与土壤含水量间的相互关系 Fig. 4 The each relation between transpiration rate, stomatic resistance of Locust, Arborvitae & Apple with soil water content |
不同土壤水分梯度的林木蒸腾速率观测结果(图 4,表 1)表明:蒸腾速率随土壤含水量变化的总趋势,可以用三次四项式描述为:y=ax3+bx2+cx+d。式中:y为蒸腾量;x为土壤含水量;a,b,c,d为拟合参数。Tr随θ的变率为:dy/dx=3ax2+2bx+c。
令dy/dx = 0,可求出Tr最高时所对应的土壤含水量,即蒸腾速率Tr峰值处的土壤含水量的临界值,侧柏为19.7%(田间持水量93.8%),刺槐和苹果分别为18.82 %(田间持水量89.6%)和16.33%(田间持水量77.8%)。
从Rs与Tr的关系上看(图 4C),随Rs的增加,Tr呈幂指数递减。但在不同的土壤含水量范围内,Rs对Tr的影响程度不同(图 4B)。整体上看,当Rs值在2~3 s·cm-1以下,土壤含水量在10%以上时,蒸腾速率可保持在较高的水平上。
2.3 叶片水分利用效率与土壤水分的关系叶片水分利用效率取决于Pn与Tr的比值,是干旱气候环境下确定栽培植物的种类、种植方式和评价其水分生产力的重要指标。在干旱环境条件下,植物水分利用效率的大小决定了植物节水能力和水分生产力水平。叶片水分利用效率与土壤水分关系的研究是确定作物不同生育期田间水分管理指标和措施的重要依据。很多研究认为,WUEL并非在水分充足时最高,而在适度的水分胁迫范围内,由于光合速率Pn随Cs的适度减小而下降的速度滞后于蒸腾速率Tr的下降速度,即由于蒸腾作用对水分胁迫的响应比光合作用敏感,蒸腾作用超前于光合作用下降,使WUEL有所提高,并且作物在适度水分亏缺时仍可获得较高的产量(山仑等,1991;徐世昌等,1995)。
根据上述研究结果,三树种的叶片水分利用效率WUEL随土壤含水量SWC的变化呈三次四项式的关系,具体变化过程近似横S形(图 5)。结果表明:在充分供水的条件下,土壤含水量15 %~25%范围内,三个树种的WUEL均出现低谷区。

|
图 5 刺槐、侧柏和苹果叶片水分利用效率与土壤含水量的关系 Fig. 5 The relation between leaf water use efficient of Locust, Arborvitae & Apple with soil water content 刺槐Locust○观测值Measured data;
—模拟值Modeled line 侧柏Arborvitae ▲观测值Measured data;—模拟值Modeled line 苹果Apple ●观测值Measured data; --模拟值Modeled line |
刺槐WUEL的第一峰值区出现在土壤含水量8%~14%左右范围,即田间持水量38%~67%,呈一定阈值变化,根据数学拟和式计算,与峰值对应的土壤含水量为13.23%,相当于田间持水量63%。
苹果峰值区对应土壤含水量为10%~15%范围,WUEL峰值对应的土壤含水量为12.38% (田间持水量59%)。
侧柏峰值区对应土壤含水量为8%~12%范围,相当于田间持水量38%~57%,WUEL峰值处于土壤含水量10.66% (田间持水量的51%)处。
3 结论综合上述光合速率、羧化效率、水分利用效率与土壤含水量关系的研究分析,可以基本确定出半干旱区集水造林适宜刺槐、侧柏和苹果生长的林地土壤水分调控标准。
刺槐净光合速率最高时的土壤含水量为17.13%,维持最高光合速率70%以上的土壤含水量最低应在10.25%以上;维持叶片水分利用效率的最佳土壤含水量为13.23%;羧化效率最高时的土壤含水量临界值为15.7%;土壤水合补偿点为4.55%。
侧柏光合速率最高时的土壤含水量为15.9%,维持70%最大光合速率的土壤含水量应在9.27 %以上;叶片水分利用效率最高时的土壤含水量为10.66%;土壤水合补偿点为3.91%。
苹果净光合速率最高的土壤含水量为16.11%,维持最大光合速率70%以上水平相对应的最低土壤含水量为9.75%;维持最高叶片水分利用效率的土壤含水量为12.38%;羧化效率最高时的土壤含水量为15.2%;土壤水合补偿点4.49%。
上述三个树种蒸腾速率所适宜的土壤含水量均在15%以上,侧柏为19.70%,刺槐为18.82%,苹果为16.33%。
根据集水造林的基本构思,干旱半干旱地区防护林的营造和经济林管理是建立在有效提高水分利用效率的基础之上,充分供水以达到最高产量不是径流林业的发展目标,更何况最高产量的获取必然伴随着牺牲水分利用效率的高昂代价。所以,维持高效蒸腾速率的土壤水分不是径流林业所期待的结果。
因此,选取维持较高光合速率所需土壤含水量的低限值,即刺槐10.25%、苹果9.75%、侧柏9.27%,选取维持最高叶片水分利用效率的土壤含水量临界值,即刺槐13.23%、苹果12.38 %和侧柏10.66%,作为集水造林林木生长适宜土壤水分环境的土壤含水量低限和高限值。这一指标可概化为刺槐10%~13%,苹果10%~12.5%,侧柏9.5%~11%。
郭柯, 董学军, 赵雨星, 等. 1996. 植物剪枝蒸腾速率变化规律的初步研究. 植物学报, 38(8): 661. |
康绍忠, 刘晓明, 王振镒. 1991. 冬小麦叶水势、气孔阻力、蒸腾速率与环境因子的关系. 灌溉排水, 10(3): 1-5. |
刘奉觉, 郑世锴编著. 1991. 杨树水分生理研究. 北京: 北京农业大学出版社, 35-40.
|
刘奉觉, 郑世锴, 巨关升, 等. 1997. 林木蒸腾耗水测算方法的比较研究. 林业科学, 33(2): 117-126. DOI:10.3321/j.issn:1001-7488.1997.02.003 |
刘奉觉. 1988. 使用稳态气孔计的几个技术问题. 林业科技通讯, (2): 31-32. |
刘昌明, 王会肖著. 1999. 土壤-作物-大气界面水分过程与节水调控. 北京: 科学出版社, 30-37.
|
邹琦.植物光合作用的气孔与非气孔限制.见: 邹琦等主编.作物抗旱生理生态研究, 北京: 中国农业科学技术出版社, 1994: 155~163
|
李吉跃. 1991. 太行山主要造林树种耐旱特性的研究(Ⅴ)—耐旱生产力. 北京林业大学学报, (增刊2): 251-264. |
王斌瑞. 1988. 晋西黄土高原主要树种凋萎湿度的研究. 北京林业大学学报, 10(4): 17-23. |
康绍忠, 刘晓明, 熊运章著. 1994. 土壤-植物-大气连续体水分传输理论及其应用. 北京: 水利电力出版社, 122-147.
|
王斌瑞, 王百田. 1996. 黄土高原径流林业. 北京: 中国林业出版社, 17-31.
|
山仑, 徐萌. 1991. 节水农业及其生理生态基础. 应用生态学报, 2(1): 70-76. DOI:10.3321/j.issn:1001-9332.1991.01.012 |
徐世昌. 1995. 水分胁迫对玉米光合性能及产量的影响. 作物学报, 2(3): 354-363. |