 2002, Vol. 38
2002, Vol. 38文章信息
- 史作民, 程瑞梅, 刘世荣, 蒋有绪, 陈宝金.
- Shi Zuomin, Cheng Ruimei, Liu Shirong, Jiang Youxu, Chen Baojin.
- 宝天曼植物群落物种多样性研究
- STUDY ON SPECIES DIVERSITY OF PLANT COMMUNITIES IN BAOTIANMAN
- 林业科学, 2002, 38(6): 17-23.
- Scientia Silvae Sinicae, 2002, 38(6): 17-23.
-
文章历史
- 收稿日期:2001-04-27
-
作者相关文章


2. 山东省教育学院 济南 250013
2. Shandong Education College Ji' nan 250013
生物群落是在一定地理区域内, 生活在同一环境下的不同种群的集合体, 其内部存在着极为复杂的相互关系。群落多样性就是指群落在组成、结构、功能和动态方面表现出的丰富多彩的差异。因此, 群落多样性是群落生态学研究, 乃至整个生态学研究中十分重要的内容。其中群落在组成和结构上表现出的多样性是认识群落的组织水平, 甚至功能状态的基础, 也是生物多样性研究中至关重要的方面。不同的植物群落在结构和功能上都存在很大的差异, 而具有不同功能作用的物种及其个体相对多度的差异是形成不同群落的基础。因此, 对于群落组织化程度的测度指标即物种多样性的研究具有重要意义(马克平, 1993; 1994;1995)。宝天曼自然保护区位于我国北亚热带向暖温带和第二级阶梯向第三级阶梯的过渡区, 特殊的地理位置和自然条件使该区植物群落类型复杂, 生物多样性相对丰富。近年来我们已对该区植物群落的区系组成、种群生态位、数量分类与排序、种间联结性等进行了分析(史作民等, 1996; 1999;2000), 本文对该区植物群落物种多样性进行了初步研究。
1 研究地区自然概况宝天曼自然保护区位于河南省西南部内乡县境内, 秦岭东段, 伏牛山南坡, 地理坐标为北纬33°25′~ 33°33′, 东经111°53′~ 112°。山体呈东南西北走向, 总面积约53.4 km2, 最高峰宝天曼海拔1 830 m。总辐射量平均为108.83 kcal·cm-2, 年均气温15.1 ℃, 1月份平均气温1.5℃, 7月份平均气温27.8 ℃。高山区无霜期160 d, 低山区无霜期227 d。年均降水量885.6 mm, 年均蒸发量991.6 mm, 年均相对湿度为68 %。土壤垂直分布比较明显, 海拔1 300m以上为山地棕壤, 800 ~ 1 300 m为山地黄棕壤, 600~ 800m为山地褐土。植被以暖温带落叶阔叶林为主, 兼有北亚热带常绿落叶阔叶混交林的特点(史作民等, 1996)。
2 研究方法 2.1 样方设置本研究主要采用样地法取样, 取样面积分别为:乔木样地20 m ×20 m, 灌木样地20 m ×20 m。每个乔木样地分成4个10 m×10 m的乔木样方, 每个乔木样地内设5个2 m ×2 m的灌木样方和5个1 m ×1 m的草本样方; 每个灌木样地内设置5个2 m ×2 m的灌木样方和5个1 m ×1 m的草本样方。调查记录每个样方内乔木的种类、数量。高度、胸径、基径、冠幅, 灌木的种类、数量、高度、盖度, 草本植物的种类、数量、平均高、盖度, 同时测定记录各样地的海拔、坡度、坡向。共设置调查了100个乔木样地, 2个灌木样地, 计有种子植物362种。全部工作于1994 -06 ~ 08和1995 -06 ~ 08完成(史作民等, 2000)。
2.2 多样性测度方法采用目前应用较为普遍的计算公式, 对研究地区植物群落物种多样性进行测度(Magurran, 1988; 谢晋阳, 1993; 马克平, 1994; 郝占庆等, 1994)。
2.2.1 丰富度指数物种丰富度指数

|
Simpson指数
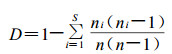
|
Shannon-Wiener指数

|
Pielou均匀度指数

|
Alatalo均匀度指数

|
式中, ni为第i种的个体数, n为所有种的个体总数, Pi为第i种的个体数ni占所有种个体总数n的比例, 即Pi =ni/n; i =1, 2, 3, ……S, S为物种数。
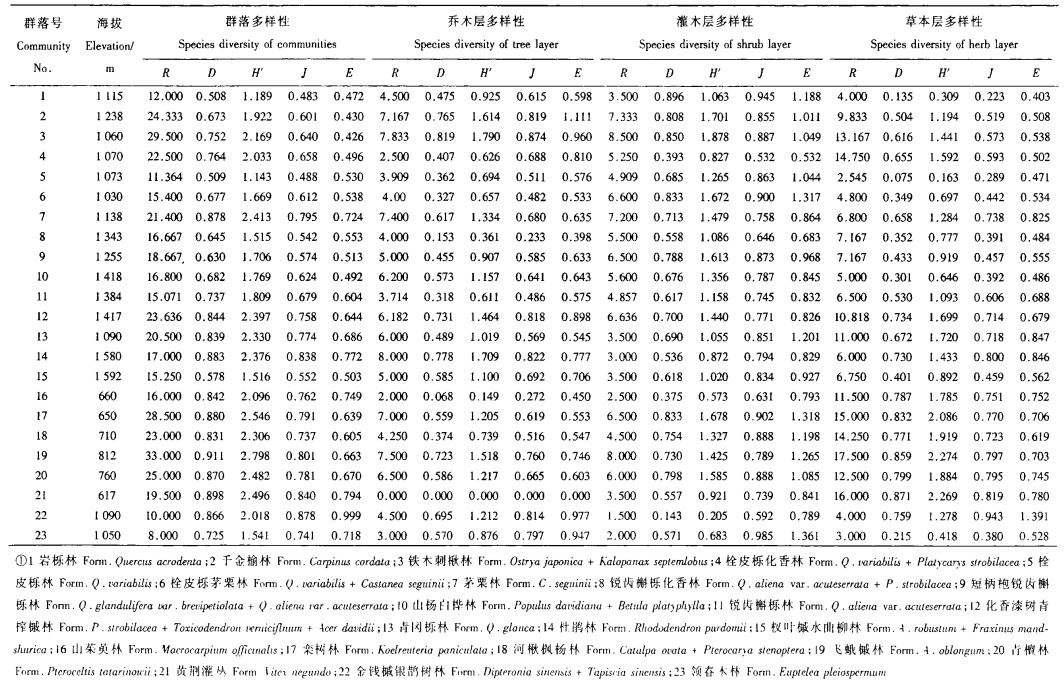
3 结果 3.1 物种多样性群落梯度上的分布由于物种多样性是一综合度量, 进行多样性分析时应将各多样性测度指标进行全面考虑。植物物种多样性(丰富度、多样性和均匀度)5种指数在群落梯度上的分布趋势基本一致(表 1)。栓皮栎林和岩栎林主要分布在向阳山坡, 群落内生境相对比较严酷, 林下灌木和草本植物相对贫乏, 领春木林和金钱槭银鹊树林主要分布在环境比较阴湿的沟谷, 林下过度阴湿的环境同样造成了灌木和草本植物种类的相对稀少, 这四种群落类型的物种丰富度和均匀度都较低, 从而导致了多样性指数较低:铁木刺楸林、化香漆树青榨槭林和飞蛾槭林由于分布在士壤和水热条件均较好的生境中, 其多样性指数较高。总之, 丰富度指数、多样性指数和均匀度指数较好地反映了不同植物群落类型在物种组成方面的差异。
|
|
植物生活型(life form)是表征群落外貌特征和垂直结构的重要指标。基于本区植物群落的结构特征, 主要讨论乔木层、灌木层和草本层物种多样性特征, 考虑到该地区层间植物种类较少, 且其功能与灌木相似, 将其以灌木计算。
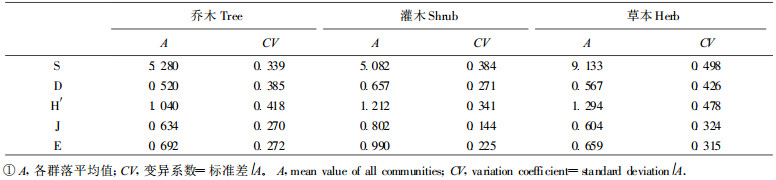
3.2.1 物种丰富度与生活型的关系除少数植物群落(栓皮栎林、栓皮栎茅栗林、茅栗林、山杨白桦林和杜鹃林)的草本层物种丰富度指数稍低于乔木层或灌木层外, 大多数植物群落的草本层物种丰富度指数明显高于乔木层和灌木层, 而各群落乔木层和灌木层物种丰富度指数没有明显的差异(表 1)。差异性检验结果也表明, 乔木和草本以及灌木和草本的物种丰富度指数间有显著差异, 乔木和灌木的物种丰富度指数差异不显著(表 2)。这种结果与北京东灵山地区温带落叶阔叶林物种多样性特点基本类似, 而与亚热带常绿阔叶林和热带地区季雨林和季节性雨林的物种多样性特点有明显的区别(黄建辉等, 1997), 即它基本上反映了本过渡区植被以温带为基调的特点。
|
|

乔木层、灌木层和草本层物种丰富度指数在群落梯度上存在一定差异, 而且有大致相同的变化趋势。乔木层:最大值为最小值的4倍, 灌木层:最大值几乎为最小值的6倍, 草本层:最大值为最小值的5倍多(表 1)。不同生活型植物物种多样性的群落间变异分析表明, 草本层物种丰富度指数在群落梯度上的变异最大, 灌木层物种丰富度指数次之, 乔木层物种丰富度指数变异最小(表 3)。
|
|
由于受物种丰富度和均匀度的双重影响, 不同生活型植物物种多样性指数在群落梯度上的分布没有明显的规律(表 1)。部分群落的灌木层物种多样性指数大于乔木层和草本层, 该结果与亚热带地区常绿阔叶林的物种多样性特点比较一致, 而有别于北京东灵山地区温带落叶阔叶林、热带季雨林及季雨林(黄建辉等, 1997), 即它从另一侧面反映了该地区植被具有一定的亚热带特征; 另一部分群落的草本层物种多样性指数大于乔木层和灌木层。相对而言, 乔木层物种多样性指数较小。差异性检验结果表明乔木和灌木物种多样性指数间有显著差异, 乔木和草本以及灌木和草本的物种多样性指数差异不显著(表 2)。
不同生活型植物物种多样性指数在群落梯度上有一定差异。乔木层物种多样性指数H′和D的最大值为最小值的12倍多; 灌木层H′的最大值为最小值的9倍多, D的最大值为最小值的6倍多; 草本层H′的最大值接近最小值的14倍, D的最大值接近最小值的12倍(表 1)。不同生活型植物物种多样性的群落间变异分析表明, 草本层和乔木层物种多样性指数在群落梯度上的变异程度明显大于灌木层(表 3)。由于H′和D的计算方法不同, H′值大于D值, 所以各生活型植物H′在群落梯度上的变异程度大于D。
3.2.3 物种均匀度与生活型的关系群落的物种均匀度是指群落中各个物种的多度或重要值的均匀程度, 它所表征的是群落观察多样性与群落种数及总个体数相同时的可能最大多样性之间的比率(岳明等, 1997)。宝天曼不同生活型植物物种均匀度指数在群落梯度上的分布没有明显的规律, 相对而言, 灌木层物种均匀度指数大于乔木层和草本层, 而乔木层和草本层物种均匀度指数除少数群落有明显不同外, 大部分群落中无明显差异(表 1)。这反映出群落的结构与组织化水平, 即乔木层和草本层物种间的重要值差异较大, 优势种表现明显; 灌木层则相反, 优势成分不明显, 常见种(common species)和稀少种(rare species)的差距较小。乔木层和草本层的物种丰富度指数差异较大, 但物种均匀度指数无明显差异, 说明这两层的多度分布比较接近。差异性检验结果表明虽然乔木层和灌木层、乔木层和草本层以及灌木层和草本层的物种均匀度指数间存在一定差异, 但其差异均不显著(表 2)。
不同生活型植物物种均匀度指数在群落梯度上有一定差异。乔木层层物种均匀度指数J的最大值接近最小值的4倍, E的最大值接近最小值的3倍; 灌木层J的最大值接近最小值的2倍, E的最大值为最小值的2.5倍多; 草本层J的最大值为最小值的4倍多, E的最大值接近最小值3.5倍。比较而言, 不同生活型植物物种均匀度指数在群落梯度上的变异程度低于物种丰富度指数和物种多样性指数。相对而言, 草本层和乔木层物种均匀度指数在群落梯度上的变异程度大于灌木层(表 3)。
3.3 群落物种多样性随海拔梯度的变化由表 1可见, 群落物种丰富度指数和物种多样性指数在海拔梯度上的变化趋势基本一致, 而物种均匀度指数的变化趋势与其存在一定差别。物种丰富度指数和物种多样性指数的较大值基本上同时出现在低海拔的栾树林和飞蛾槭林, 中等海拔的铁木刺楸林, 以及高海拔的化香漆树青榨槭林等植物群落; 较小值同时出现在中等海拔的栓皮栎林、岩栎林、领春木林和金钱槭银鹊树林等植物群落。物种均匀度指数的较大值基本上同时出现在中等海拔的金钱槭银鹊树林, 低海拔的黄荆灌丛, 以及高海拔的杜鹃林等植物群落; 但物种均匀度指数J和E的较小值出现在不同的植物群落, J的较小值出现在中等海拔的岩栎林和栓皮栎林等植物群落, E的较小值出现在中等海拔的千金榆林和铁木刺楸林等植物群落。
3.4 不同生活型植物物种多样性在海拔梯度上的分布 3.4.1 不同生活型植物物种丰富度指数在海拔梯度上的分布不同生活型植物物种丰富度指数在海拔梯度上的分布规律存在一定差异。除海拔1 600 m附近的杜鹃林具有最高的乔木层物种丰富度指数和较小的灌木层物种丰富度指数外, 乔木层和灌木层物种丰富度指数的较大值基本上出现在中等海拔的茅栗林、铁木刺楸林、千金榆林和低海拔的飞蛾槭林等植物群落。草本层物种丰富度指数的较大值出现在低海拔的飞蛾槭林、栾树林、河楸枫杨林和黄荆灌丛等植物群落(表 1)。分布于低海拔的山茱萸林由于是人工或半人工林而表现出最低的乔木层物种丰富度指数。分布于中等海拔的金钱槭银鹊树林和领春木林由于林下过度荫湿的生境和较高的乔木层郁闭度, 导致灌木层物种丰富度指数和草本层物种丰富度指数较小。
3.4.2 不同生活型植物物种多样性指数在海拔梯度上的分布不同生活型植物物种多样性指数在海拔梯度上的分布及其相互关系没有明显的规律。乔木层物种多样性指数的较大值出现在中等海拔的铁木刺楸林、千金榆林和高海拔的杜鹃林等植物群落, 较小值出现在低海拔的山茱萸林以及高海拔的锐齿栎化香林和锐齿栎林等植物群落。灌木层物种多样性指数较大值出现在中等海拔的铁木刺楸林、千金榆林和低海拔的栾树林等植物群落, 较小值出现在低海拔的山茱萸林以及中等海拔的金钱槭银鹊树林和领春木林等植物群落。草本层物种多样性指数较大值出现在低海拔的栾树林、黄荆灌丛和飞蛾槭林等植物群落, 较小值出现在中等海拔的栓皮栎林、岩栎林和领春木林等植物群落(表 1)。
3.4.3 不同生活型植物物种均匀度指数在海拔梯度上的分布与物种多样性指数相似, 不同生活型植物物种均匀度指数在海拔梯度上的分布及其相互关系没有明显的规律。乔木层物种均匀度指数的较大值出现在中等海拔的青冈栎林、铁木刺楸林和低海拔的河楸枫杨林, 较小值出现在中等海拔的锐齿栎林和低海拔的山茱萸林。灌木层物种均匀度指数的较大值出现在中等海拔的栓皮栎茅栗林、领春木林和低海拔的栾树林, 较小值出现在中等海拔的栓皮栎化香林、金钱槭银鹊树林和高海拔的锐齿栎化香林。草本层物种均匀度指数较大值出现在中等海拔的金钱槭银鹊树林和低海拔的黄荆灌丛, 较小值出现在中等海拔的岩栎林和栓皮栎林(表 1)。
由表 1还可以看出, 灌木层物种均匀度指数高于乔木层和草本层, 即海拔梯度上灌木层的多度分布相对乔木层和草本层而言比较一致。灌木层物种均匀度指数基本上表现出随海拔升高而降低的趋势, 这表明高海拔区灌木层的常见种和稀少种较低海拔区明显, 低海拔区灌木层的优势种不明显。
4 讨论与结论 4.1 讨论山地植被群落物种多样性及其随海拔梯度的变化规律一直是生态学家感兴趣的问题, 开展的研究比较多, 但研究结果不尽一致。通过对有关山地植被群落物种多样性研究工作的综述, 贺金生等(1997)将山地植被群落物种多样性指数随海拔高度的变化模式划分为5种:①植物群落物种多样性指数与海拔高度负相关, 即随海拔高度的升高, 植物群落物种多样性指数降低; ②植物群落物种多样性指数在中等海拔高度最大, 也有学者称之为“中间高度膨胀”(Whittaker et al., 1975;Peek, 1978); ③植物群落物种多样性指数在中等海拔高度较低; ④植物群落物种多样性指数与海拔高度正相关, 即随海拔高度的升高, 植物群落物种多样性增加; ⑤植物群落物种多样性指数与海拔高度无关。由此可见, 山地植被群落物种多样性指数随海拔梯度的变化没有统一的规律, 二者的相互关系比较复杂。事实上, 山地植被分布区域的环境条件、山地的相对高度、人为干扰程度, 不同海拔高度的群落类型、群落的发育阶段、群落分布区的坡位、坡度和坡向以及群落内土壤厚度、有机质含量和水分条件等都有可能对植物群落物种多样性指数在海拔梯度上的分布产生影响(Peek, 1978; Wilson et al., 1990;Itow, 1991; Baruch, 1984; 谢晋阳等; 黄建辉, 1994; 黄建辉等, 1997; 高贤明等, 1998)。
宝天曼保护区最高海拔仅1 830 m, 为中山区。我们将该山地海拔800 m以下的部分称为低海拔区, 海拔800 ~ 1 200 m的部分称为中等海拔区, 海拔1 200 m以上的部分称为高海拔区。依据此划分范围, 宝天曼保护区植物群落物种丰富度指数和物种多样性指数随海拔梯度的变化在一定程度上呈现出高海拔和低海拔区偏高、中等海拔高度偏低的趋势。不同海拔范围人类活动的干扰状况、群落类型、群落演替阶段、群落小生境以及水热条件的组合等是影响群落物种多样性指数的重要因子。此外, 草本层物种丰富度和物种多样性指数大于乔木层和灌木层这一特征导致了草本层植物的变化对不同海拔植物群落物种多样性指数的影响较大。低海拔区人类活动相对频繁, 对该区各类型植物群落存在一定程度的干扰。特别是目前还不太严重的森林放牧活动导致了以群落的郁闭度稍低、林下阳光相对充足和枯枝落叶层较薄等为主要表现形式的群落生境的变化, 从而为灌木层和草本层植物创造了较为优越的生存环境, 结果群落内林下植物较为丰富, 群落物种多样性较高。中等海拔区人类活动较少, 植物群落郁闭度较高, 林下生境或过度阴湿而不利于灌木和草本植物的生长, 或相对干旱积累了较厚的枯枝落叶层而影响草本植物的生长, 从而导致该区植物物种多样性偏低。另外, 该区植物群落多属于森林皆伐后形成的次生群落, 成熟水平相对较低, 基本处于群落演替的中期阶段, 结构与功能还没有达到最优水平, 这也是导致植物物种多样性偏低的因素之一。高海拔区几乎没有人类干扰, 部分植物群落成熟水平较高, 基本处于群落演替的顶级阶段, 结构复杂, 功能优化, 群落内物种之间的生态位重叠较大, 相互依存的能力较高。另外, 该区水热条件组合最好, 相对湿润的林下生境以及分解较快的较薄的枯枝落叶层均有利于林下植物的生长。上述两个主要因素使得该区的植物群落具有偏高的物种多样性特征。
4.2 结论物种丰富度指数、多样性指数和均匀度指数在群落梯度上的分布趋势基本一致, 较好地反映了不同植物群落类型在物种组成方面的差异。
多数植物群落的草本层物种丰富度指数明显高于乔木层和灌木层, 而乔木层和灌木层物种丰富度指数没有明显的差异。草本层物种丰富度指数在群落梯度上的变异最大, 灌木层次之, 乔木层变异最小。乔木层物种多样性指数小于灌木层和草本层。草本层和乔木层物种多样性指数在群落梯度上的变异程度明显大于灌木层。灌木层物种均匀度指数大于乔木层和草本层, 而乔木层和草本层均匀度指数在大部分群落中无明显差异。草本层和乔木层均匀度指数在群落梯度上的变异程度大于灌木层。
植物物种丰富度指数、物种多样性指数的较大值分布在高海拔和低海拔区, 较小值多分布在中等海拔高度。植物物种均匀度指数的较小值多分布在中等海拔高度, 但其较大值在各海拔高度都有分布。
草本层植物物种丰富度指数和物种多样性指数的较高值多分布在低海拔区, 较小值多分布在中等海拔高度。乔木层和灌木层的各种指数以及草本层的均匀度指数在海拔梯度上的分布规律不明显。
马克平. 1993. 试论生物多样性的概念. 生物多样性, 1(1): 20-22. DOI:10.3321/j.issn:1005-0094.1993.01.006 |
马克平. 1994. 生物群落多样性的测度方法Ⅰα多样性的测度方法(上). 生物多样性, 2(3): 162-168. DOI:10.3321/j.issn:1005-0094.1994.03.007 |
马克平, 黄建辉, 于顺利, 等. 1995. 北京东灵山地区植物群落多样性的研究Ⅱ丰富度、均匀度和物种多样性指数. 生态学报, 15(3): 268-277. DOI:10.3321/j.issn:1000-0933.1995.03.006 |
史作民, 刘世荣, 王正用. 1996. 河南宝天曼种子植物区系特征. 西北植物学报, 16(3): 329-335. DOI:10.3321/j.issn:1000-4025.1996.03.020 |
史作民, 程瑞梅, 刘世荣. 1999. 宝天曼落叶阔叶林种群生态位特征. 应用生态学报, 10(3): 265-269. DOI:10.3321/j.issn:1001-9332.1999.03.003 |
史作民, 刘世荣, 程瑞梅, 等. 2000. 河南宝天曼植物群落数量分类与排序. 林业科学, 36(6): 20-27. DOI:10.3321/j.issn:1001-7488.2000.06.003 |
岳明, 周虹霞. 1997. 太白山北坡落叶阔叶林物种多样性特征. 云南植物研究, 19(2): 171-176. |
贺金生, 陈伟烈. 1997. 陆地植物群落物种多样性的梯度变化特征. 生态学报, 17(1): 91-99. DOI:10.3321/j.issn:1000-0933.1997.01.014 |
郝占庆, 陶大立, 赵士洞. 1994. 长白山北坡阔叶红松林及其次生白桦林高等植物物种多样性比较. 应用生态学报, 5(1): 16-23. DOI:10.3321/j.issn:1001-9332.1994.01.013 |
高贤明, 陈灵芝. 1998. 北京山区辽东栎(Quercus liaotungensis)群落物种多样性的研究. 植物生态学报, 22(1): 23-32. DOI:10.3321/j.issn:1005-264X.1998.01.003 |
黄建辉. 1994. 物种多样性的空间格局及其形成机制初探. 生物多样性, 2(2): 103-107. DOI:10.3321/j.issn:1005-0094.1994.02.008 |
黄建辉, 高贤明, 马克平, 等. 1997. 地带性森林群落物种多样性的比较研究. 生态学报, 17(6): 611-618. |
谢晋阳.物种多样性指数与物种多度分布.见林金安主编.植物科学综论.哈尔滨: 东北林业大学出版社, 1993: 222~233
|
谢晋阳, 陈灵芝. 1994. 暖温带落叶阔叶林的物种多样性特征. 生态学报, 14(4): 337-344. DOI:10.3321/j.issn:1000-0933.1994.04.012 |
Baruch Z. 1984. Ordination and classification of vegetation along an altitudinal gradient in the Venezuelan paramos. Vegetatio, 55: 115-126. |
Itow S. 1991. Species turnover and diversity patterns along an elevation broad-leaved forest coenocline. Journal of Vegetation Science, 2: 477-484. DOI:10.2307/3236029 |
Mangurran A E. 1988. Ecological diversity and its measurement. Princeton: Princeton University Press, 1-179.
|
Peek R K. 1978. Forest vegetation of the Colorado Front Range:Pattern of species diversity. Vegetatio, 37: 65-78. DOI:10.1007/BF00126830 |
Whittaker R H, Niering W A. 1975. Vegetation of the Sant Catalina Mountains, Arizona :V.Biomass, production, and diversity along the elevation gradient. Ecology, 56: 771-790. DOI:10.2307/1936291 |
Wilson J B, Lee W G, Mark A F. 1990. Species diversity in relation to ultramafic substrate and to altitude in southwestern New Zealand. Vegetatio, 86: 15-20. DOI:10.1007/BF00045131 |