 2002, Vol. 38
2002, Vol. 38文章信息
- 吴若菁.
- Wu Ruojing.
- 天然马尾松群体遗传结构的研究
- GENETIC DIVERSITY OF PROTEIN IN ALCOHOL FOR PINUS MASSONIANA FROM NATURAL POPULATION
- 林业科学, 2002, 38(5): 160-165.
- Scientia Silvae Sinicae, 2002, 38(5): 160-165.
-
文章历史
- 收稿日期:2001-03-13
-
作者相关文章


醇溶蛋白是种子的贮藏蛋白,在对麦类植物的醇溶蛋白研究中,发现醇溶蛋白的电泳谱带的带纹多少及组合方式完全受基因型所控制,几乎不受环境的影响,可以构成品种的指纹(Drapar,1987;Zillman et al.,1979).因而几年来,国内外已将醇溶蛋白电泳分析应用到遗传育种、种子鉴定、资源保存利用以及物种起源、演化分类研究等方面(颜启传等,1992;傅宾孝等,1993;吴若菁等,1997;2000;杨瑞武等,2000;王志容等,2000;李名旺等,2000)。
马尾松(Pinus massoniana)广泛分布于我国南方,是主要的造纸和用材林树种,也是我国荒山绿化和营建生态保护林的先锋树种。天然马尾松林在长期进化的因素作用下,形成了一定的遗传结构和较丰富的遗传变异。为此,许多研究者采用各种研究方法来了解马尾松天然群体在不同生长环境、不同生长状况下的遗传结构、遗传多样性、繁育系统和地理变异等。在研究中应用最多的是同工酶分析技术。研究结果发现,马尾松群体同工酶存在遗传多样性(葛颂等,1987;解荷峰,1990)。但马尾松种子贮藏蛋白是否也存在遗传多样性,在不同生长环境下其遗传结构将发生怎样的变化,则未见报道。为此,本试验首次利用天然马尾松林木的种子胚乳进行醇溶蛋白电泳分析,以研究马尾松天然群体遗传结构的组成及变异情况,以便为马尾松遗传育种、资源保存利用、系统演化提供基础资料和科学依据。
1 材料和方法 1.1 材料材料取自福建省漳平市五一林场卓宅工区的天然马尾松成熟林,经度117.5°E,纬度25.2°N,林分面积7.33 hm2,300株·hm-2,采种树间距为5倍树高,分株采种,每株10个球果,共采集60株,分株净种,4 ℃贮藏种子。
1.2 样品制作浸种24 h,剥取胚乳,每个胚乳加样品提取液0.5 mL,研磨成浆,室温浸提过夜。12 000 r·min-1离心10 min,取上清液备用。(样品提取液为2-巯基乙醇1 mL,2-氯乙醇20 mL,尿素18.0 g,甲基绿0.05 g,混合后用蒸馏水定容至100 mL)。
1.3 凝胶和电泳凝胶缓冲液用冰乙酸20 mL,甘氨酸1.0 g,蒸馏水定容至1 000 mL。凝胶液用甲叉双丙烯酰胺0.577 g,丙烯酰胺14.423 g,尿素6.0 g,抗坏血酸0.1 g,硫酸亚铁0.000 5 g混合后用凝胶缓冲液100 mL溶解。
电极缓冲液用冰乙酸4 mL,甘氨酸0.8 g,蒸馏水定容至1 000 mL。分离胶制备取凝胶液25 mL,加入0.6%H2O2 0.09 mL,10%过硫酸胺0.025 mL,TEMED 0.075mL,混合均匀,灌板,每个样品槽加20 μL样品液,恒压230 V,4 ℃电泳50 min。
1.4 固定和染色染色液用考马斯亮兰R250 1.0 g溶于100 mL乙醇中。电泳完毕后,将凝胶取下先用10%三氯乙酸200 mL固定0.5 h,再加入染色液30 mL,染色过夜,拍照。
1.5 基因型分析每单株分析6~10粒种子的胚乳,确定基因型,当基因位点为异质时,其等位基因分离比例应为1:1,采用经过Yatas′矫正的χ2进行检验。
1.6 等位基因频率检测与统计检测了428粒种子的胚乳谱带,按已确定各位点的各等位基因图谱表型直接计算。
1.7 多态位点的标准本文采用的多态位点的标准是指第2个等位基因的频率超过0.01(Nei,1973)。
1.8 统计分析分析群体遗传结构的几个主要衡量指标,按王中仁先生(1996)的方法进行。
2 结果与讨论马尾松含有二倍体的胚和单倍体的胚乳,在实验中可直接利用单倍体胚乳的醇溶蛋白电泳图谱来分析其是否遵循孟德尔的分离规律。通过χ2检验可以较容易地确定电泳谱带所代表的基因位点及位点内所具有的等位基因。当基因位点为异质时,则该位点的两个等位基因分离比例应为1:1。因此,在这种分析条件下,把杂合基因型错定为同质基因型的概率(1/2)8-1≈0.008。
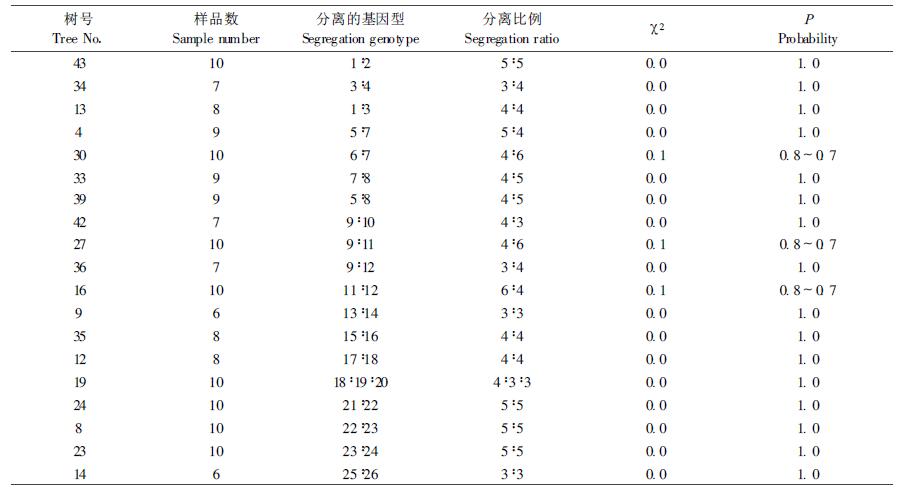
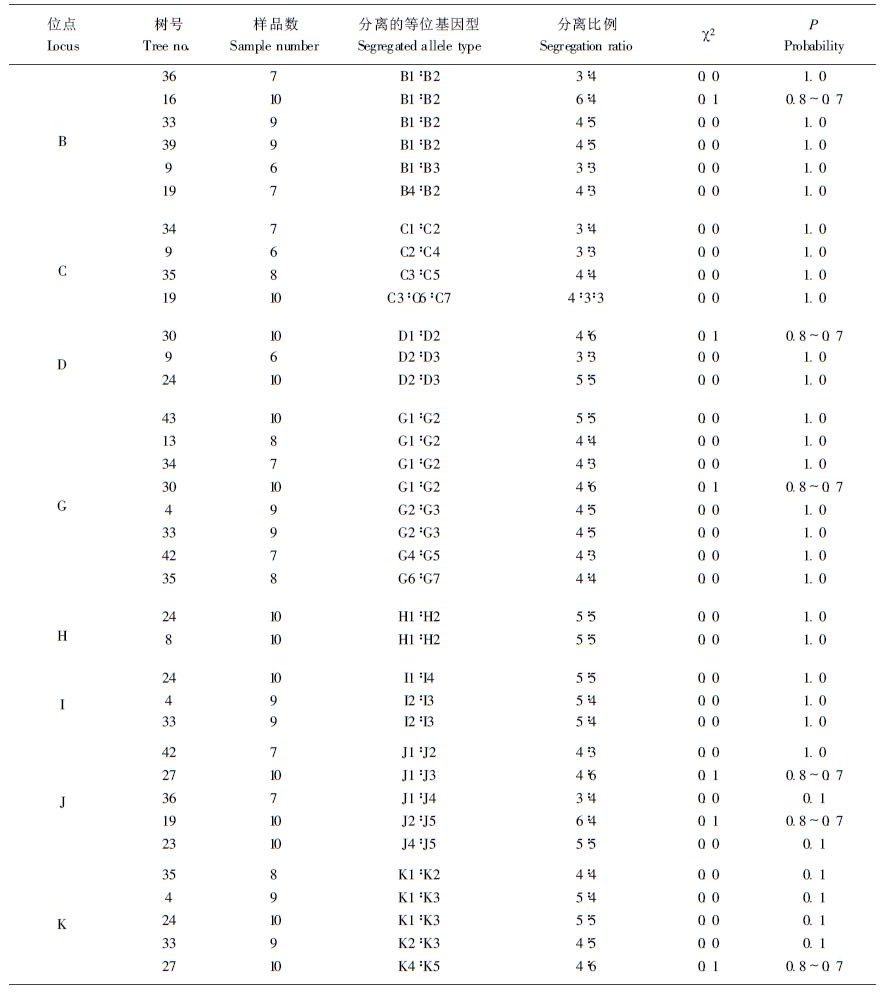
本研究以单倍体的胚乳为材料,分株测定,共检测60株,每株检测6~10粒种子的胚乳,共获得428个有效样品,发现该天然马尾松林中存在有26种类型的醇溶蛋白图谱,群体内的杂合单株的醇溶蛋白图谱的表型分离,经χ2检验,符合孟德尔1:1的分离比(见图 1、表 1)。此外,还发现一种少见的醇溶蛋白图谱类型,该类型具C7带,在G位点具2条染色特别深的带,其相对迁移率与G位点的7种带均不同,而且,还具I4、J5带,这2条带着色也很深。这种特异类型的图谱出现频率仅占所分析总样品的0.6%,而且不符合正常的孟德尔分离规律。在符合分离规律的26种醇溶蛋白图谱中,共获得11个位点,其中A、E、F 3个位点为单态位点,其余8个为多态位点。这8个多态位点所具有的等位基因数2~7个;有3个高活性位点区,分别为B、D、G基因位点。对群体内各位点的杂合单株中两个等位基因的分离比进行χ2检验均符合1:1孟德尔分离比率(见表 2、图 2)。说明确是属于同一位点上不同等位基因所控制的。

|
图 1 马尾松醇溶蛋白电泳谱类型 Fig. 1 Electrophoretic patterns of protein patterns in alcohol for Pinus massoniana |
|
|
|
|

|
图 2 马尾松醇溶蛋白不同位点等位基因图谱 Fig. 2 Alleles pattens of various loci for protein in alcohol in Pinus massoniana |
天然林木群体内的遗传多样性,既是长期进化的产物,又是反映该种群进化的潜能指标。本研究对马尾松天然群体遗传结构衡量指标的分析结果为:多态位点百分数P=72.7%,平均每个位点基因平均数A=3.636,平均每个位点等位基因有效数目Ne=2.663,平均每个位点的预期杂合度He=0.552,平均每个位点的实际杂合度Ho=0.231,详见(表 3)。从这些遗传变异的主要衡量指标可以看出,本研究所分析的马尾松天然群体的遗体多样性水平是较高的,这主要是因为马尾松为较原始的裸子植物,分布范围较广,又是绿化荒山的先锋树种,其生存所处的环境条件要求树木必须具有较复杂的遗传基础,较宽的适应性。因此,马尾松天然群体在长期的进化过程中经受着各种不同的选择压力,从而产生了丰富的遗传多样性。据研究资料报道松属植物中同工酶的遗传变异变化很大,从脂松(P.reasiocsis)的P=0 (Fowler et al.,1977)到欧洲赤松的P=100%(Hamrick et al.,1979),不同物种显示了不同的变异水平,与其它已研究过的松属植物相比(Fowler et al.,1977;Hamrick et al.,1979;1982;Kim et al.,1995),马尾松同工酶研究中发现其P为68.06%~77.78%,居于松属树种中较高水平,本试验对马尾松醇溶蛋白分析的结果,多态位点百分数P=72.7%与同工酶研究的结果十分接近。但是,马尾松醇溶蛋白的A=3.636、Ne=2.663,He=0.552,Ho=0.231这些指标高于同工酶分析结果(解荷峰,1990),说明马尾松醇溶蛋白的遗传变异比同工酶的3个酶系统(GOT,MDH,GDH)更大。
|
|
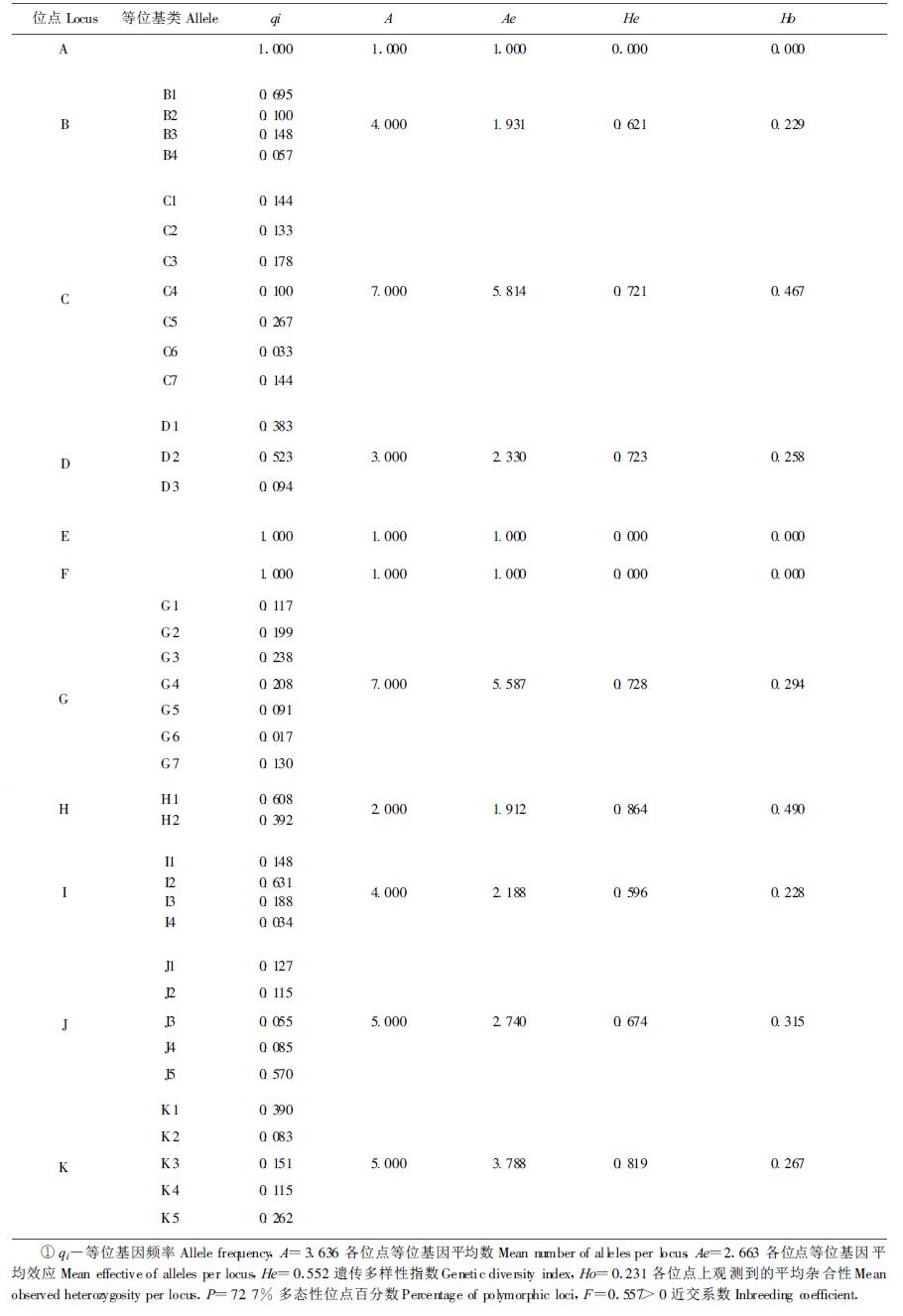
分析的结果还发现,该马尾松自然群体的固定指数F=0.557 >0,偏离了哈迪一温伯格平衡,显示群体中纯合体过量。其原因是马尾松虽然是风媒异交种,而且本研究所取材料的群体分布面积也较大,但是该群体中个体密度太小,在这种条件下,就不能产生充分的随机交配,易出现遗传漂移,并使自交或近亲交配比率增高,使得群体纯合性增加,杂合性下降,出现偏离群体遗传平衡的现象。
综上所述,马尾松种子胚乳的醇溶蛋白电泳分析可作为一种新的衡量指标,较好地反映群体内的遗传组成及群体的遗传动态变化情况,用于指导育种实践。
傅宾孝, 于光华, 王乐凯, 等. 1993. 小麦醇溶蛋白电泳分析的新方法. 作物学报, 19(2): 185-187. DOI:10.3321/j.issn:0496-3490.1993.02.014 |
葛颂, 黄敏仁, 许农. 1987. 马尾松GOT、LDH和MDH同工酶的遗传方式和连锁关系. 遗传学报, 14(6): 428-435. |
解荷峰. 1990. 马尾松天然群体的遗体学结构. 南京林业大学学报, 14(4): 36-43. |
李名旺, 於丙军, 顾德兴, 等. 2000. 半夏属的花粉粒和醇溶蛋白比较. 武汉植物学研究, 18(3): 250-252. DOI:10.3969/j.issn.2095-0837.2000.03.015 |
王志容, 魏育明, 兰香锦, 等. 2000. 四川三个专用小麦的贮藏蛋白分析. 四川农业大学学报, 18(1): 18-20. DOI:10.3969/j.issn.1000-2650.2000.01.006 |
王中仁. 1996. 植物等位酶分析. 北京: 科学出版社, 145-154.
|
吴若菁, 苏红蓉, 危彝斌, 等. 1997. 马尾松与黑松种子醇溶蛋白鉴别的研究. 福建林学院学报, 17(4): 293-296. |
吴若菁, 赖文胜, 程朝阳, 等. 2000. 火炬松等5种松树的醇溶蛋白图谱研究. 福建林学院学报, 20(1): 46-48. DOI:10.3969/j.issn.1001-389X.2000.01.013 |
杨瑞武, 周永红, 郑有良. 2000. 几种特异小麦的醇溶蛋白电泳分析. 四川农业大学学报, 18(1): 15-17. DOI:10.3969/j.issn.1000-2650.2000.01.005 |
杨瑞武, 周永红, 郑有良. 2000. 披碱草属的醇溶蛋白研究. 四川农业大学学报, 18(1): 11-14. DOI:10.3969/j.issn.1000-2650.2000.01.004 |
颜启传, 黄亚军, 徐媛. 1992. 试用ISTA推荐的种子醇溶蛋白电泳方法鉴定大麦和小麦品种. 作物学报, 18(1): 61-67. DOI:10.3321/j.issn:0496-3490.1992.01.008 |
Drapar S R. 1987. ISTA variety committee report of the working group for biochemical tests for cultivar identification 1983-1986. Seed Sci & L Technol, 15: 431-434. |
Fowler D P, Morris R W. 1977. Genetic diversity in red Pine: evidenci for low genetic heterozygosity. Can J For Res, 7: 343-347. DOI:10.1139/x77-043 |
Hamrick J L, et al. 1979. Relationship between life history characteristics and eletroetically detedtable genetic variation in plant. Ann Rev Ecol Syst, 10: 173-200. DOI:10.1146/annurev.es.10.110179.001133 |
Hamrick J L. 1982. Plant population genetic. Amer J Bot, 69: 1685-1693. DOI:10.1002/j.1537-2197.1982.tb13421.x |
Kim Zing Suk, Sook Woo Lee. 1995. Genetic diversity of three native Pinus in Korea, SPB. Acadmic Publish, 211-218.
|
Nei M. 1973. Analysis of gene diversity in subdivided populations. Proc Nat Acad Sci USA, 70(12): 3321-3323. DOI:10.1073/pnas.70.12.3321 |
Zillman R R, Bushuk W. 1979. Wheat cultivar identification by gliadin electrophoregrams. Ⅱ Effects of envionmental and experimental factors on the gliadin electrophoregrams. Can J Plant Sci, 59: 281-286. DOI:10.4141/cjps79-047 |
Zillman R R, Bushuk W. 1979. Wheat cultivar identification by gliadin electrophoregrams. Ⅲ. Catalogue of electrophoregrams fomulas of canadian wheat cultivars. Can J Plant Sci, 59: 287-288. DOI:10.4141/cjps79-048 |