 2002, Vol. 38
2002, Vol. 38文章信息
- 池玉杰, 于钢.
- Chi Yujie, Yu Gang.
- 6种木材白腐菌对山杨材木质素分解能力的研究
- STUDY ON THE DEGRADING ABILITY TO DAVID POPLAR LIGNIN OF SIX SPECIES OF WOOD WHITE-ROT FUNGI
- 林业科学, 2002, 38(5): 115-120.
- Scientia Silvae Sinicae, 2002, 38(5): 115-120.
-
文章历史
- 收稿日期:2001-10-15
-
作者相关文章


由于不同的木材腐朽菌生理特性不同, 所分泌的酶及酶的活性各不相同, 因此, 不同的腐朽菌所分解木材的各种成分及相对速度各不相同(Buswell, 1987)。有些木腐菌只能分解纤维素和木质素二者之一; 有些木腐菌则可同时分解二者; 还有些菌类能以二者之一为主要分解对象, 兼或稍具分解另一物质。所以在自然界中, 木材腐朽的类型是很复杂的。如木质细胞壁中木质素被分解利用时, 仅留下纤维素, 朽材较健康材色浅, 呈白色, 就叫白腐。朽材不易粉碎解体, 且富于弹性, 木材质地变为纤维状或海绵状, 这样的白腐木材有可能用于造纸。
山杨(Populus davidiana), 又称中国山杨, 属白杨派, 分布于北纬25°~ 53°、东经100°~ 130°范围内, 常形成天然次生林, 与桦木类混生或成纯林, 为采伐迹地和火烧迹地天然更新的先锋树种, 是中国山区造林先锋树种之一。从造纸的特定材性指标上看, 山杨具有较好的造纸纤维形态, 纤维较长, 不易断裂; 综纤维素含量较高(80 %), 木质素含量较低(20 %), 制浆得率高; 材色洁白而易漂白, 材质轻软而易于脱除木质素, 易得高白度浆, 因此是造纸的良好材料。在自然状态下山杨活立木却极易染心材白腐病, 山杨心材白腐, 也叫阔叶树心材白腐, 广泛发生在山杨、白桦、青杨、色木槭、蒙古栎、核桃楸等许多种阔叶树上, 尤其山杨林、青杨林和白桦林最多, 遍布世界各地, 在我国东北、西北、西南和华北各地林区普遍发生。引起山杨心材白腐的木腐菌为火木层孔菌(Phellius igniarius)。火木层孔菌在野外状态下危害树干心材, 发病初期心材变褐色, 逐渐褪为白色, 并在其周围产生黑色线纹, 最后朽材变软, 不碎不裂, 形成典型的白色海绵状腐朽。在大风雨天, 树干常从腐朽部折断, 露出雪白松软的朽材。这种自然状态下腐朽后的木材用于造纸的情况还有待深入地研究, 因此作者选择了山杨材和火木层孔菌, 以探讨木材白腐菌对木材与木质素的生物降解机制。
作者曾在以前的研究中测定了东北林区60余种木腐菌对青杨、白桦和红松的木材分解能力, 得出了木腐菌对青杨、白桦和红松的木材分解能力排序表(池玉杰, 2001), 但木腐菌对木质素分解能力的研究在国内还未见报道(Tanesaka, 1993)。木质素是植物木质化组织的重要成分, 其化学结构相当复杂, 而且性状十分稳定, 但是在自然界中还是存在具木质素分解能力的微生物。千万年来, 森林中的枯枝落叶、倒木等未堆积成山即是证明, 只是不同的微生物对木质素的分解程度不同。根据迄今的研究结果, 在所有具分解木质素能力的微生物中, 只有引起木材白色腐朽的真菌被确切地证明了能够彻底地分解木质素, 能把复杂的木质素高分子降解成CO2和H2O(Kirk, 1984; Paterson, 1985)。近年来, 微生物在制浆造纸中的作用被重视起来, 提出了生物法制浆造纸的研究课题。木材造纸的一大难题是脱除木材中的木质素, 但会对水资源造成污染。如果利用微生物对木质素的降解作用脱除木材中的木质素, 就从根本上解决了造纸工业的环境污染问题。无疑地, 研究与采用无污染或少污染的制浆与漂白新技术是我国造纸业的紧迫任务, 生物制浆将是造纸工业的发展方向。有希望可以利用的是高效选择性分解木质素的白腐菌, 将木材中的木质素分解后剩下纤维素以用于造纸。
在木材中, 木质素的贮量仅次于纤维素, 但是长期以来对木质素的利用还不够深入, 不少场合如在造纸业中却作为废弃物加以排除, 不但带来严重的环境污染, 也浪费资源。因此, 提高木质素的利用效率、开发木质素的利用途径, 是解决资源、能源、环境问题的一个重要措施。从理论上讲, 控制性地对木质素进行生物降解, 可以由这一复杂的高聚物生产出现在从石油中制取的各种有用的苯酚类物质(Crawfod, 1981; Tai, 1990), 如香草酸、二甲基硫醚、丁香酸、苯酚等。这种办法如果成功, 这将为当前与未来的化学品生产开辟新的原料来源。为此, 国外有些人做了这种以生物法从木质素中制取有用化学品的探索性研究。利用白腐菌对木质素的分解作用, 是一种很有希望的方法。因此作者选择了火木层孔菌及另外5种木材分解能力较强的阔叶树上的白腐菌:粗毛盖菌(Funalia gallica)、三色革裥菌(Lenzites tricolor)、冬拟多孔菌(Polyporellus brumalis)、偏肿拟栓菌(Psendotrametes gibbosa)和血红密孔菌(Pycnoporus sanguineus), 研究了它们对山杨材木质素的分解能力。目的为山杨材生物制浆造纸提供应用基础理论, 同时也可为木质素合理的生物转化为有用的化学品、生物漂白、酶处理、防止机械浆的返黄、废水治理、纤维素酶解糖化的微生物前处理等提供相关的借鉴研究, 以期在生产实践上减轻环境污染并充分利用木质素资源。
1 材料与方法 1.1 试验材料 1.1.1 6种木材白腐菌培养物的制备将在东北林区采集到的有生命力的木腐菌子实体, 用组织分离法得到菌种。在无菌条件下, 将在斜面培养基上生长7 d的木腐菌菌种, 切取直径为5 mm的同质等量菌丝块, 分别接入到PDA平板培养基的中间部应, 每一菌种多个重复, 接种后的培养基置于25 ℃恒温箱中培养7 ~ 15 d, 使菌种长满平板或基本长满平板。
1.1.2 采样在冬季采伐期间进行采样。选择东北林业大学帽儿山实验林场20 ~ 30 a生山杨人工林的未腐朽材。本批样品是2000-11-29采自帽儿山林场新垦事业区7林班20 ~ 30 a生小径级山杨。木材样品采回后及时晾干, 以免受杂菌的侵染。
1.1.3 木材样品的制备根据国家标准———木材天然耐腐性实验室试验方法(GB/T13942.1-93), 将野外采回的山杨木材, 晾干到一定程度后, 锯成2 cm×2 cm ×1 cm小片。
1.2 试验方法 1.2.1 木片样品受白腐菌腐朽试验将一定数量的木片样品, 置于105 ℃烘箱中烘至恒重, 在高压灭菌锅中灭菌(0.12MPa下灭菌30 min), 试样含水率达到40 %~ 60 %, 在无菌条件下将木片放入接种10 d已长满菌丝的平板培养基内, 然后将培养皿放入25 ℃恒温箱中培养使木片受菌侵染, 并保持一定湿度。对于每一菌种, 山杨木片样品分别被腐朽一定时期(为40 d, 60 d, 80 d和120 d; 不同的时间所用木片样品分别是40 d —120 ~ 140块, 60 d —160 ~ 200块, 80 d —200 ~ 260块, 120 d —240 ~ 320块)后, 取出附带菌丝体的小块, 用毛刷和水冲洗干净, 除去表面菌丝, 然后放入105 ℃烘箱中烘至恒重, 根据受菌侵染前后木片样品的重量变化计算受菌分解后木片样品重量减少百分率, 并记录受菌侵染后木片的颜色变化(国家标准, GB/T 1394.1-93), 并按1.2.2测定受菌分解后木片的木质素含量变化。根据木片样品重量变化和木质素的含量变化计算木片样品中综纤维素的含量变化。
1.2.2 山杨木片样品木质素含量的测定通过测定受白腐菌分解前后的木片样品中木质素的含量变化可知木质素被分解的程度, 木质素减少百分率与木片样品重量减少百分率的比率可表明木质素被分解的难易程度。按照国家标准一造纸原料分析用试样的采取(GB/T 2677.1-93)和造纸原料酸不溶木素含量的测定(GB/T 2677.8-94)进行分析侵染前后山杨材木片样品的木质素含量。本项试验测定了6种白腐菌分别分解山杨材木片样品40 d、60 d、80 d和120 d时的木质素含量及未腐朽时山杨木材木质素的含量, 共得到25项木质素含量百分率的测试数据, 最后得到木材样品中酸不溶木质素的含量
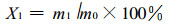
|
式中:m1为烘干后的酸不溶木质素的质量(g); m0为绝干试样质量(g)。
2 结果与讨论山杨木片样品受菌侵染前后木质素的含量变化结果见表 1。根据山杨材受菌侵染后重量损失百分率和木质素分解百分率可得出综纤维素分解百分率。

|
|
|
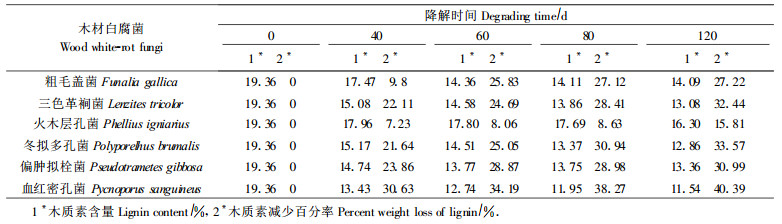
设腐朽前木片试样的绝干重为W1, 腐朽后木片试样的绝干重为W2, 那么腐朽前综纤维素含量= W1 -W1 ×木质素含量百分率= W1 ×(1 -19.36 %)=0.8064W1; 腐朽后综纤维素含量=W2 -W2 ×腐朽后木质素含量百分率= W2(1 -腐朽后木质素含量百分率)。
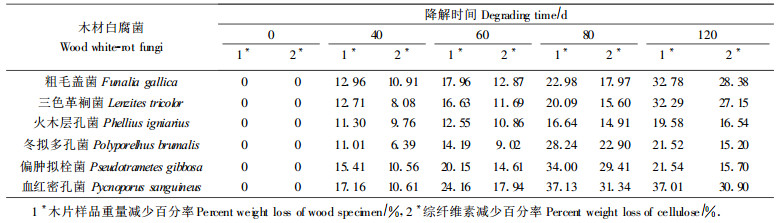
山杨木片样品受菌侵染后的木材分解百分率和综纤维素的含量变化结果见表 2。
|
|
由于木腐菌的生理特性不同, 所分泌的酶及酶的活性各不相同, 因此, 不同的木腐菌分解木材的各种成分及相对速度就各不相同, 不同的白腐菌对于相同的木质纤维基质也有不同的中间代谢产物和途径。从表 1可以看出, 6种白腐菌对山杨木质素的分解能力不同。木片受白腐菌分解后的木质素减少百分率可作为评价白腐菌木质素分解能力的指标, 木质素减少百分率愈高, 说明白腐菌对木质素的分解能力愈强; 而木质素减少百分率愈低, 说明白腐菌对木质素的分解能力愈弱。根据木质素的减少百分率可以得出这6种白腐菌对山杨木质素的分解能力依次为血红密孔菌、偏肿拟栓菌、冬拟多孔菌、三色革裥菌、粗毛盖菌和火木层孔菌。
2.2 6种白腐菌对山杨木材综纤维素的分解能力和最佳白腐菌的确定在分析白腐菌对山杨材木片木质素分解能力的同时, 也要考虑白腐菌对纤维素的分解, 因为我们要寻求的是对木质素的分解能力强、尽可能少分解或不分解纤维素的选择性极高的最适于生物制浆用的白腐菌, 因此也要确定木质素被分解到什么程度时纤维素被分解的程度最低, 从而可保持最大的制浆得率。根据木片受菌分解前后的重量变化和木质素的含量变化可计算出综纤维素的含量变化。从表 2可以看出, 冬拟多孔菌在分解同样多木质素时, 综纤维素的分解率最低。因此, 在本项试验的6种白腐菌中冬拟多孔菌就是较多的分解木质素而较少的分解纤维素的较好的木材白腐菌。
2.3 木片受菌侵染时间的确定在自然状态下, 相对于其他的降解物来说, 微生物对处于固态环境中的木材和木质素的降解是一个较为缓慢的过程。在实验室内模拟自然状态的木材分解过程, 因此木材受菌侵染试验分别选择了40 d、60 d、80 d和120 d后测定木腐菌对木材和木质素的分解能力, 如果少于40 d, 木腐菌对木材和木质素的分解百分率就很低(Tanesaka, 1993), 为了要在应用的角度进一步深入考虑工艺上的可能性, 迅速缩短微生物对木片的分解周期, 就必须要找到加速微生物对木质素分解的方法, 是否可以选择复合菌种的液体发酵法还有待进一步深入研究。可以考虑木片在受菌类分解过程中, 在发酵罐中加入一些打磨木片的材料, 不断地搅拌, 以打磨木片和分散菌体, 从而加速分解过程。另外制浆原料木片要尽量薄一些, 以易于菌类的分解。能否找到能催化微生物对木质素分解的“催化剂”, 从而迅速加快这种酶解反应, 也是一个可以考虑的方法。从表 1可以看出, 多数菌种在40 d时对木质素的分解就己达到一定程度, 而且随着时间的延长木质素被分解的程度变化不十分显著, 因此通过延长时间以获得对木质素较多的分解是不合理的, 也是不经济的。在将来的生产实际操作中, 可根据试验结果, 确定一个最佳的木片受菌侵染时间, 这时已分解足量的木质素同时纤维素被分解的程度最低, 从而在纸浆生产中可保持最大的制浆得率, 而生产周期又不至于很长。
2.4 受菌侵染后木片变黄的原因分析所有受菌侵染后的木片都从白色变成了乳黄色、淡黄色, 受火木层孔菌侵染的木片还出现了黑线纹, 而变色部分主要在腐朽木片的表层受白腐菌分解的部分。这种现象产生的原因是木质素在分解过程中产生一些有色物质, 这些有色物质主要来自木质素的氧化产物, 即一些发色基因, 木质素的发色基因主要有3种类型, 即木质素的醌型结构、邻苯二酚的重金属螯合物及与结构单元的共轭结构(陈嘉翔等, 1990); 另外, 木材中的低分子有机物和单宁等也有着色作用。正是这些具有着色作用物质, 是使受菌侵染的木片变黄的原因, 这也是制浆过程中木浆变黄的原因。若能减少在木片受菌类发酵过程中木质素降解产物的氧化作用, 就可能降低纸浆的颜色。根据这一试验结果, 可以考虑在未来的生物制浆过程中应尽量减少木质素的氧化作用, 因此需要筛选厌氧发酵木片的木材白腐菌。
2.5 火木层孔菌不是最佳的白腐菌根据试验结果可以看出, 火木层孔菌不是最佳的白腐菌。在实验室条件下, 火木层孔菌的生长速度缓慢, 它的木材分解能力较弱, 木质素的分解能力也较弱, 而在野外这种木腐菌能广泛分布, 普遍生长在山杨、白桦、青杨、色木槭、蒙古栎、核桃楸等许多种阔叶树上, 引起心材腐朽的主要原因是由于火木层孔菌是具有忍耐特点的活立木心材上的腐朽菌, 它可以在其它微生物较少存在的活立木心材这一具有生存压力的特殊生境中长期生活。在活立木心材中, 最明显的压力因素是缺少易同化的营养基质, 存在着抑制性物质如单宁、树脂和芳香油等, 以及缺少氧气(程东升, 1993)。树木的心材与边材的明显区别之一就是心材中含有大量的沉积物质, 这类废弃物质是树木生命活动过程中产生的, 它们沉积在心材细胞中, 减少了心材的空隙, 并且有显著毒性, 因此一般来说心材比边材更抗腐。此外, 这些沉积物还有使心材更为稳定的趋势。火木层孔菌在培养基上的生长速度非常缓慢, 井缺少同其它微生物的竞争力, 在自然状态下多年生。这些性状都表明它的生存策略就是能够长期忍耐活立木心材这个特殊生境中的压力因素而得以生存与发展。由于具有忍耐或抵抗能力, 所以在不利因素存在的情况下, 火木层孔菌的生存能力明显大于其它非忍耐对策微生物的生存能力, 并对材积的破坏力极大。
3 结论6种白腐菌对山杨木材木质素的分解能力不同, 根据木质素的减少百分率可以得出其分解能力依次为血红密孔菌、偏肿拟栓菌、冬拟多孔菌、三色革裥菌、粗毛盖菌和火木层孔菌。
如果考虑白腐菌对纤维素的分解, 在本项试验的6种白腐菌中冬拟多孔菌是较多的分解木质素而较少的分解纤维素的较好的木材白腐菌。
在将来的生产实际中, 可确定一个最佳的木片受菌侵染时间, 这时已分解足量的木质素同时纤维素被分解的程度最低, 从而在纸浆生产中可保持最大的制浆得率, 而生产周期又不至于很长。
所有受菌侵染后的木片均从白色变成了乳黄色、淡黄色。木质素在分解过程中, 都能产生一些有色物质, 这些有色物质主要来自木质素的氧化产物, 这些具有着色作用的物质, 是使受菌侵染的木片变黄的原因, 这也是制浆过程中木浆变黄的原因。
陈嘉翔, 李元禄, 张志芬, 等. 1990. 制浆原理与工程. 北京: 轻工业出版社.
|
程东升. 1993. 森林微生物学. 哈尔滨: 东北林业大学出版社.
|
池玉杰. 2001. 东北林区64种木材腐朽菌木材分解能力的研究. 林业科学, 37(5): 107-112. DOI:10.3321/j.issn:1001-7488.2001.05.019 |
中华人民共和国国家标准.木材天然耐久性试验方法, 木材天然耐腐性实验室试验方法.GB/T 13942, 1~93
|
中华人民共和国国家标准.造纸原料酸不容木素含量的测定.GB/T 2677, 8~94
|
中华人民共和国国家标准.造纸原料分析用试样的采取.GB/T 2677, 1~93
|
Buswell J A. 1987. Odier E Lignin biodegradation. CRC Crit.Rev.Biotechnol, (6): 1-60. |
Crawfod. 1981. Microbial conversions of lignin to useful chemicals using a lignin-degrading Streptomyces.3rd symp.On biotechnology in energy production and conservation. Gatlinbury: 275-291. |
Kirk T K.Degradation of lignin.In: Microbial degradation of organic compounds.M arcel Dekker Inc, 1984, 399~437
|
Paterson A, Lundquist K. 1985. Radical breakdown of lignin. Nature, 361(6029): 575-576. |
Tai D S, Terazawa M, Chen C L. 1990. Lignin biodegradation products from birch wood decayed by Phanerochaete chrysosporinm. Holzfoschung, 44(4): 257-262. DOI:10.1515/hfsg.1990.44.4.257 |
Tanesaka E, Masuda H, Kinugawa K. 1993. Wood degrading ability of basidiomycetes that are wood decomposers.Litte decomposers, or mycorrhizal symbionts. Mycologia, 85(3): 347-354. DOI:10.1080/00275514.1993.12026283 |