 2002, Vol. 38
2002, Vol. 38文章信息
- 程水源, 王燕, 李俊凯, 顾曼如, 束怀瑞.
- Cheng Shuiyuan, Wang Yan, Li Junkai, Gu Manru, Shu Huairui.
- 银杏叶黄酮类化合物合成代谢规律的研究
- STUDY ON THE SYNTHESIS AND METABOLISM OF THE FLAVONOIDS IN GINKGO BILOBA LEAF
- 林业科学, 2002, 38(5): 60-63.
- Scientia Silvae Sinicae, 2002, 38(5): 60-63.
-
文章历史
- 收稿日期:2001-10-25
-
作者相关文章


2. 山东农业大学园艺学院 泰山 271018
2. Shandong Agricultural University Taishan 271018
黄酮类化合物在植物体内合成代谢的起初源为光合产物, 合成的前体是简单酚类, 酚类物质的合成是从苯丙氨酸解氨酶催化苯丙氨酸脱氨反应开始的, 但在酚类物质向黄酮类化合物的转化过程中同时存在多种成份的合成途径, 至少还包括木质素、生物碱、角质、栓质等(程水源等, 2000; 鞠志国等, 1992)。对花青苷而言, 它是黄酮类化合物进一步转化代谢的产物。还有, 可溶性蛋白质合成也是从苯丙氨酸合成酶催化合成苯丙氨酸开始的, 即可溶性蛋白质合成与黄酮类化合物合成是从苯丙氨酸开始走两条完全不同的途径, 存在竞争相同底物的关系(郗荣庭, 1997; Harborne, 1976)。银杏因富含黄酮类化合物而具有重要的药用价值, 但对其黄酮类化合物与其代谢转化产物、前体物质及竞争物的研究, 至今仍是空白1)。本试验选银杏叶丛枝叶片为试材, 探讨黄酮类化合物合成代谢与木质素、简单酚类、花青苷和可溶性蛋白质之间的关系, 旨在了解黄酮类化合物在银杏叶片中的合成代谢规律, 并为制定适宜采叶期和调控银杏叶黄酮类化合物提供理论依据。
1)程水源. 影响银杏叶黄酮类化合物形成的主要因子及调控技术的研究. 博士学位论文. 泰山: 山东农业大学, 2001. 5~ 11:43.
1 材料与方法 1.1 选材、取样及预处理试材选于湖北农学院银杏科技园7 a生“家佛手”, 树势中等, 生长结果正常, 株行距2 m ×3 m。在1998年, 从7月4日开始, 每隔15 d左右取叶样测定黄酮类化合物、简单酚类、木质素、花青苷、可溶性蛋白质含量, 摸索测定方法和了解各成份变化的大致规律1)。在1999年, 从5月4日开始, 每隔15 d左右在36株银杏树的叶丛枝上随机采叶100片, 取样到11月29日止。采样后迅速将叶片洗净、晾干, 经105℃杀酶20 min, 65 ℃烘干至恒重、粉碎, 干燥器贮存待测。花青苷与可溶性蛋白质含量采用鲜样测定。
1.2 测定方法与数据图表处理黄酮类化合物测定采用程水源等(2001a)的方法, 木质素测定采用鞠志国等(1992)的方法, 二者单位均为干重百分含量(% DW)。花青苷测定采用赵宗方等(1992)的方法。可溶性蛋白质测定采用Bradford(1976)的方法。简单酚类测定1):称取叶干样0.15 ~ 0.2 g, 用1 %HCl的甲醇溶液研磨提取并定容至25 mL, 然后放在冰箱中静置24 h, 再离心, 上清液用V-530分光光度仪于280 nm、530 nm、600 nm处测定吸光值。以A280 -(A530 -A600)×63%=0.1作为一个简单酚类单位(U), 以U·(g DW)-1表示简单酚类含量。
试验处理及数据测定均重复3次, 平均值表示结果, 数据统计与制图采用软件DPS、Microsoft Excel。
2 结果与分析 2.1 银杏叶黄酮类化合物含量变化及与有关成份的关系从表 1可知:银杏叶黄酮类化合物含量有2次明显的含量高峰, 第1次在7月4日, 第2次在11月1日, 含量值分别为0.808 2 %、1.062 5 %。相比较而言, 第2次高峰值比第1次提高了19.09 %, 持续时间也较第1次高峰长, 即从9月底开始一直维持在较高的含量水平。在出现含量最高峰的11月, 前半个月含量的波动仅为1.5 %, 整个11月叶黄酮类化合物含量从最高峰1.062 5 %下降到1.030 6%, 也只下降了3个百分点, 这说明叶黄酮类化合物含量在第2个合成高峰期比较稳定, 在此期间内采收叶片比较适宜。
|
|
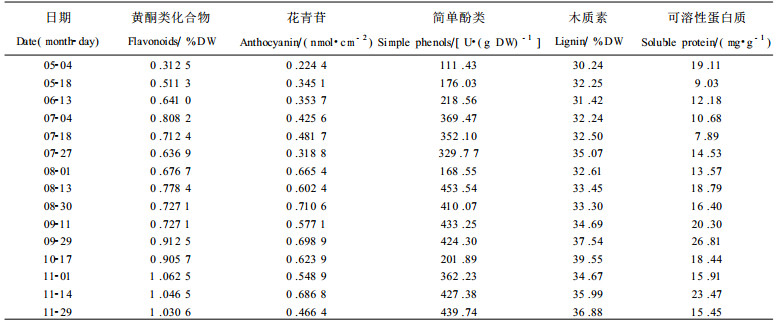
花青苷含量变化与叶黄酮类化合物有类似趋势, 但波动幅度大、次数多。用含量相对增长速率(%)比较二者(图 1):前、后期花青苷小于叶黄酮类化合物, 中期大于叶黄酮类化合物(前、中、后期的时间段分别为7月底以前、8月初至9月底、10月初以后)。这说明简单酚类向黄酮类化合物的转化过程中, 以合成强度为衡量指标, 在前、后期以合成叶黄酮类化合物为主, 而中期则以花青苷为主。这也说明黄酮类化合物与花青苷在植物体内的合成代谢具时间分布特征(原永兵等, 1995; 鞠志国等, 1993)。

|
图 1 银杏叶黄酮类化合物、花青苷含量相对变化 Fig. 1 Relative changes of flavonoids and anthocyanin contents in Ginkgo biloba leaves ①黄酮类化合物Flavonoids, ②花青苷Anthocyanin. |
在前期, 简单酚类与黄酮类化合物同步上升, 于7月初达到第1次高峰后下降, 8月1日达到最低值。此后, 简单酚类含量又急剧上升, 8月中旬至9月底是简单酚类的合成高峰期, 其含量处于较高的、较稳定的水平。8月初其含量下降与木质素的合成有关, 9月底后其含量的陡降与黄酮类化合物和木质素同时大量合成有关(表 1)。
木质素含量第1次高峰在7月底(35.076 %), 第2次在10月中旬(39.55 %), 与黄酮类化合物含量峰相比, 第1次峰滞后, 第2次峰超前。在前、中期和行进落叶期, 二者变化趋势相反, 但在中后期(1999-08-01 ~ 1999-10-17)二者呈极显著正相关, r =0.9168**。这说明来源于相同前体底物的黄酮类化合物与木质素之间, 除了有相互制约关系外, 还在一定时期内可以相互促进, 这可能与它们本身在一定阶段的合成强度和前体物质简单酚类含量有关1)。综合分析黄酮类化合物、木质素与简单酚类之间的关系, 有回归方程Y(黄酮类化合物)=-1.023 7 +0.000 8X1(简单酚类)+0.044 9X2(木质素), 调整后 R2 =0.640 4, 经F检验, 回归模型达极显著水平。从上可知, 木质素单位变量(ΔX2)对黄酮类化合物的影响比简单酚类(ΔX1)大得多, 这为调控银杏黄酮类化合物含量提供了有益的启示。
在前、后期, 黄酮类化合物含量低时, 可溶性蛋白质含量较高, 反之亦然。但在7月18日~ 9月29日, 二者呈显著正相关, r =0.935 4**, 回归方程为:Y (可溶性蛋白质)=47.063 5X(黄酮类化合物)- 16.574 2, 调整后 R2 =0.843 7, 经F检验, 回归模型达极显著水平。这也反应了叶中可溶性蛋白质与黄酮类化合物之间的关系具有阶段性的特点。
2.2 叶黄酮类化合物与相关成份的相关和偏相关分析从表 2可知:叶黄酮类化合物与花青苷、简单酚类、木质素呈显著正相关, 与可溶性蛋白质之间相关达不到显著水平, 但在偏相关分析中[参与偏相关分析的其它成分有:含水量、可溶性糖、淀粉、可滴定酸(程水源等, 2001b)], 叶黄酮类化合物仅与可溶性蛋白质呈极显著负相关, 与花青苷、简单酚类及木质素之间无显著相关关系, 相关与偏相关分析表明了不同的试验结论。
|
|
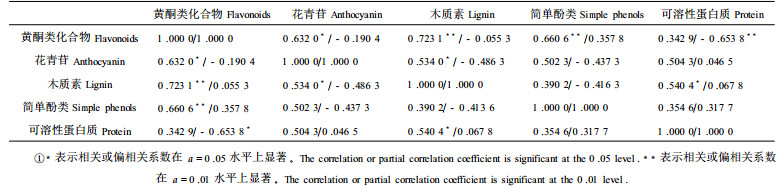
简单酚类作为黄酮类化合物合成的前体物质(程水源等, 2000; 鞠志国等, 1992; 1993), 其含量的提高, 伴随叶黄酮类化合物合成的加速和含量的提高。但当以酚类物质作底物的次生代谢产物如黄酮类化合物、木质素及花青苷等在10月中旬大量合成时, 导致简单酚类含量下降, 当黄酮类化合物等合成速率下降或含量由于分解转化而下降时, 又可见简单酚类含量上升, 这有力地说明了它们之间存在着直接的、简单的依存关系。但偏相关分析表明二者相关不显著, 这又说明在简单酚类向黄酮类化合物转化过程中由于有多种产物同时生成(程水源等, 2000; 鞠志国等, 1992; 1993), 从而导致二者关系的复杂性。
前期银杏叶片中有木质素合成, 且维持在较稳定水平。鞠志国等(1993)报道:苹果幼果皮几乎无木质素的合成, 与本试验结果不一致, 这可能与树种不同, 尤其与研究器官不同有关。幼果皮不合成木质素, 但随细胞膨大, 木质素逐渐形成(鞠志国等, 1993; Vance et al., 1980), 而本试验在5月份测定银杏叶片木质素含量时, 叶片处在相当于果实细胞膨大后的发育阶段。用不同时间段来分析木质素与叶黄酮类化合物的关系表明:前期和行进落叶期二者变化相反, 在中后期(8月初~ 10月中旬), 二者含量变化趋势相近, 这说明在前期及行进落叶期简单酚类主要用于次生代谢产物如叶黄酮类化合物合成, 但在中后期, 由于简单酚类的大量合成与积累, 虽然有叶黄酮类化合物与木质素同时合成, 但看不出因前体底物不够而相互制约的现象。不过, 可以看出由于它们竞争同一前体物质而使简单酚类含量在9月底至10月初表现出大幅度下降。简单相关分析表明:木质素与叶黄酮类化合物之间呈极显著正相关, 但偏相关分析结果是:二者无显著正相关, 甚至偏相关系数还变成负值。这说明了木质素和黄酮类化合物含量的变化只是随简单酚类含量的变化而作出的相应反应, 并非二者之间存在真实的显著性关系:互相促进, 相反, 正是由于竞争同一前体底物而表现出互相制约的趋势。作为黄酮类化合物代谢转化产物的花青苷, 与黄酮类化合物的关系类似于木质素, 只是花青苷对黄酮类化合物合成的影响程度比木质素更大一些, 这与花青苷与黄酮类化合物在合成途径中密切程度有关(程水源等, 2000; 鞠志国等, 1992)。
叶中可溶性蛋白质也是黄酮类化合物的竞争物, 二者之间相关达不到显著水平, 但偏相关分析呈极显著负相关。这说明在排除其它因素影响的情况下, 二者之间有真实负相关关系, 这可以从它们的合成途径中得到解释(程水源等, 2000; 郗荣庭, 1997; Harborne, 1976)。前人研究叶片中含有过量的氮, 尤其是采叶前期含有过量的氮或者可溶性蛋白质, 不利于黄酮类化合物的形成。在施肥试验中, 后期给土壤大量施氮, 叶黄酮类化合物含量下降, 进一步支持了上述观点(Max, 1990)。但当叶片光合能力强, 前体底物含量较高, 虽然二者合成速率上升, 但总体表现较为平稳时, 它们之间有可能并不表现出明显的反向关系, 本试验结果证明了这一推断。7月中~ 9月底, 它们之间呈显著正相关, 正好说明了它们之间由于竞争同一底物而表现的相互促进关系, 即通过竞争, 促进光合产物的转化, 表现为二者含量的提高。合成可溶性蛋白质与合成黄酮类化合物是走两条不同的途径, 蛋白质比简单酚类、花青苷和木质素(后三者与叶黄酮类化合物合成是走同一途径)对叶黄酮类化合物合成的影响大得多。因此, 在调控银杏叶黄酮类化合物的研究中, 叶中可溶性蛋白质的作用与地位不容忽视。
程水源, 顾曼如, 束怀瑞. 2000. 银杏叶黄酮类化合物研究进展. 林业科学, 36(6): 110-115. DOI:10.3321/j.issn:1001-7488.2000.06.019 |
程水源, 王燕, 李俊凯, 等. 2001a. 银杏叶片黄酮类化合物含量变化与分布规律的研究. 园艺学报, (4): 353-355. |
程水源, 王燕, 李俊凯, 等. 2001b. 银杏叶类黄酮合成代谢与叶中有关成份关系的研究. 华中农业大学学报, 20(5): 474-477. |
鞠志国, 刘成连, 原永兵, 等. 1992. 苹果果皮中酚类物质合成规律的研究. 莱阳农学院学报, 9(3): 222-225. |
鞠志国, 刘成连, 原永兵, 等. 1993. 莱阳茌梨酚类物质合成的调节及其对果实品质的影响. 中国农业科学, 26(4): 44-48. |
郗荣庭主编. 果树栽培学总论. 第三版. 北京: 中国农业出版社, 1997, 88
|
原永兵, 刘成连, 鞠志国等. 苹果果皮红色形成的机制. 见: 园艺学年评. 北京: 科学出版社, 1995, 121~ 132
|
赵宗方, 谢嘉宝, 吴桂清, 等. 1992. 富士苹果果皮花青素的相关因素分析. 果树科学, 9(3): 134-147. |
Bradford M W. 1976. A rapid and sensitive method for the quantitation of microgram quantities protein utilizing the principle of protein-dye binging. Anal Biochem, 72: 248-254. DOI:10.1016/0003-2697(76)90527-3 |
Harborne J B. Biochemical systhematics of flavonoids. In: Hardborne J B, Mabry T J, Mabry H (eds). The Flavonoids. London: Academic press, 1976, 1056~ 1095
|
Max C S. 1990. External control of anthocyanin formation in apple. Scientia Horticulturae, 42: 181-218. DOI:10.1016/0304-4238(90)90082-P |
Vance C P, Krik T K, Sherwood R T. 1980. Lignifications of lignin mechanism of disease resistance. Ann Rev Phytopathol, 18: 259-288. DOI:10.1146/annurev.py.18.090180.001355 |