 2002, Vol. 38
2002, Vol. 38文章信息
- 昝启杰, 王勇军, 廖宝文, 郑德璋, 黄立南.
- Zan Qijie, Wang Yongjun, Liao Baowen, Zheng Dezhang, Huang Linan.
- 木榄种源引种深圳湾后幼苗生理生态研究
- A STUDY ON THE PHYSIOLOGICAL-ECOLOGY OF BRUGUIERA GYMNORHIZA SEEDLINGS AFTER INTRODUCED TO SHENZHEN BAY FROM DIFFERENT AREAS
- 林业科学, 2002, 38(5): 47-53.
- Scientia Silvae Sinicae, 2002, 38(5): 47-53.
-
文章历史
- 收稿日期:2000-02-14
-
作者相关文章


2. 中国林业科学研究院热带林业研究所 广州 510520;
3. 中山大学环境科学系 广州 510275
2. The Research Institute of Tropical Forestry, CAF Guangzhou 510520;
3. Department of Environmental Science, Zhongshan University Guangzhou 510275
红树林在我国仅天然分布于海南、广西、广东、福建、台湾等省区沿海滩涂, 且大部分集中在海南和两广地区, 红树林面积已从20世纪50年代的4×104 hm2锐减至目前的不足2×104 hm2, 占不到世界红树林面积的0.1% (郑德璋等, 1995)。为加强我国红树林资源的保护和发展, 20世纪90年代以来, 红树林引种造林研究日益受到重视。近年来人口快速增长, 工业化、城市化迅猛发展, 深圳湾红树林湿地面积大大缩小, 一些物种生存压力愈来愈大, 如红树植物木榄(Bruguiera gymnorhiza)在深圳湾将有消亡的危险。为此, “九五”国家科技攻关专题拟将优良木榄种源引种至深圳湾。本研究将不同木榄种源引种到深圳湾后, 对其幼苗形态、生长和生理生态进行研究, 分析各种源的生长差异及在深圳湾生境中的生理生态反应, 为筛选适宜在深圳湾生长的优良种源提供理论依据。
1 试验地及种源地自然概况在广东内伶仃福田国家级自然保护区红树林区域西侧海滩上, 选择秋茄+桐花树人工林的林窗作为海滩苗床, 苗床所在地位于深圳湾的东北岸, 属于南亚热带季风气候, 干湿季节交替明显, 全年主导风为东南风和东北风, 但本区东南方向有香港大雾山阻挡, 减弱了强风的袭击, 成为红树林生长的理想地方。苗床土壤基质为花岗岩及砂页岩, 地带性土壤为砖红壤, 红树林发展土壤大多淤泥浓厚, 育苗土壤为酸性硫酸土, 有效N、P和速效K分别为42.1、40.7和838.1 mg·kg-1 (廖宝文等, 1996)。
木榄种源分别采自海南三亚、琼山、文昌, 广西防城, 雷州海康, 廉江高桥及深圳本地, 试验地及各种源地自然条件见表 1。采种地群落为近海岸高潮带上的木榄群落, 高3~5 m, 郁闭度为0.7~0.85。
|
|
1997年5月下旬至6月上旬, 从海南三亚、琼山、文昌, 广西防城, 廉江高桥, 雷州海康及深圳福田本地的天然木榄母树上, 随机摘取墨绿色成熟木榄胚轴, 随即去掉萼管, 挑选完好无损、长短粗细一致的胚轴, 立即带到深圳作试验材料。
先用0.3%~0.5%的高锰酸钾溶液对苗床海泥进行消毒, 再用直径8 cm、深12 cm的育苗袋装满海泥, 且保证不同种源的育苗袋海泥基质相同, 把7个不同来源的木榄胚轴插植在育苗袋中(深度约为胚轴长度的2/3), 放在海滩苗床上培育, 随机区组排列, 每组2行, 每行30个育苗袋, 3次重复(如图 1)。

|
图 1 木榄胚轴培育设计图 Fig. 1 Planting pattern of Mangrove B.gymnorhiza Hypocotyl W:海南文昌的种源Provenance fromWenchang, Hainan, S:海南三亚的种源Provenance from Shanya, Hainan, Q:海南琼山的种源Provenance fromQiongshan, Hainan, Y:广西防城的种源Provenance fromFangcheng, Guangxi, G:廉江高桥的种源Provenance fromGaoqiao, Lianjiang, H:雷州海康的种源Provenance fromHaikang, Leizhou, F:深圳福田的种源Provenance fromFutian, Shenzhen. |
幼苗满1龄时, 测定每株高度(胚轴高度除外)和基部直径(测量胚轴上部近地面茎径, 以便排除原胚轴粗细对测定结果的影响), 统计出幼苗平均株高和基径及每个植株叶片总数量(不计子叶), 并随机测量50片成熟叶片的长度和宽度, 再用计算纸法(王伯荪等, 1996)测定叶面积。
2.3 生理指标检测叶片含水量的测定用称重法(朱广廉等, 1990)。自由水含量用阿贝氏折射仪法(朱广廉等, 1990), 将含水量减去自由水而求得束缚水。叶绿素a、b及总量的测定, 取各种源胚轴发育的木榄幼苗成熟叶片(下同), 用分光光度计法(朱广廉等, 1990), 据Lambert-Beer定律计算; 用酸性茚三酮法(朱广廉等, 1983), 测定成熟叶片游离脯氨酸的含量; 用高锰酸钾滴定法(黄学林等, 1990)测定过氧化氢酶活性; 用电导仪法(朱广廉等, 1990)测定叶片相对电导率。叶片光合作用、蒸腾作用及气孔导度的测定, 用4台美制Li-6200便携式光合作用测定仪, 在9:00、12:00、16:00测定木榄不离体叶片的净光合速率、蒸腾速率及气孔导度(同一时刻, 福田、海康、高桥、防城种源与文昌、琼山、三亚种源的测定时差小于2 min)。
2.4 数据分析与综合评价将各种源引种深圳湾后的一龄幼苗的生长及生理生态指标观测数据, 用Statgraphics Statistical Graphics System软件进行方差分析和多重比较分析。应用综合评定方法(顾万春, 1986)将幼苗生长及生理生态观测指标综合起来, 评定引种到深圳的木榄种源品质的优劣。
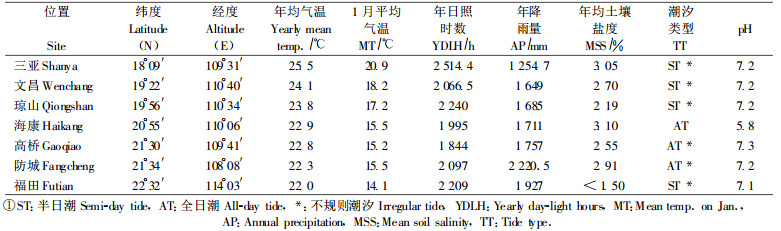
3 结果与分析 3.1 幼苗高度和基径表 2显示, 除文昌种源外的5个外来木榄种源的幼苗存活率均高于福田本地种源, 文昌种源存活率最低, 可能因为胚轴采摘后放置了近20 d才带到深圳培育, 影响了胚轴萌发和早期生长; 从株高和基径看, 高桥、三亚、琼山种源生长最好, 海康种源生长次之, 福田、防城和文昌种源生长较差; F值表明, 7个种源的存活率和地径生长都存在极显著差异, 而高度的差异不显著。从表 1可知, 各种源地环境条件差别很大, 各种源长期生活在不同的环境中, 一般都会形成不同的生态适应性, 当引种到相同生境后, 它们的生长表现出差异性, 这可能是各种源对深圳福田生境的适应差异, 也可能是各种源存在一定的生态遗传差异。
|
|
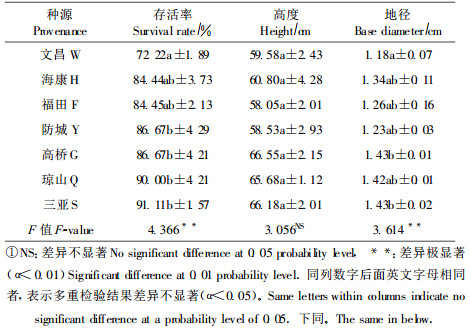
不同木榄种源的幼苗分枝数量、叶片大小等存在差异(表 3)。海康、三亚、琼山种源的分枝数明显多于福田本地种源, 而广西防城种源的分枝数最少; 琼山、高桥种源幼苗成熟叶片长、宽明显大于其它5个种源; 叶面积以琼山种源最大, 高桥种源次之, 分别比福田本地种源的大25.3%和16.2%, 而文昌种源的叶面积只有本地种源的80.4%;方差分析和F值结果表明, 各种源之间的每株叶片数量和叶面积存在极显著差异。文昌种源幼苗, 叶片明显短而窄, 叶片面积也最小, 仅有45.1 cm2, 并且幼苗叶片和海莲的叶片几乎相同。调查文昌种源采集地发现, 文昌的木榄种群和海莲种群混合在一起生长, 因此, 该地木榄极有可能不纯, 是与海莲的杂交种。
|
|
不同木榄种源在深圳福田培育后幼苗叶片及分枝的差异性, 一方面可能是物种对新环境的适应差异, 另一方面可能是各种源在原来特定的生境中, 形成的遗传性状差异。这有待作进一步的遗传分析, 并对照原产地的种源原有特性作分析研究后确定。
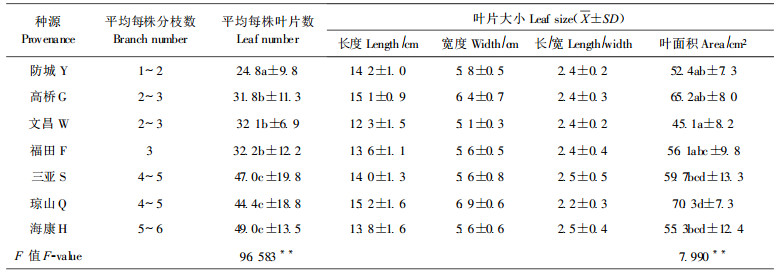
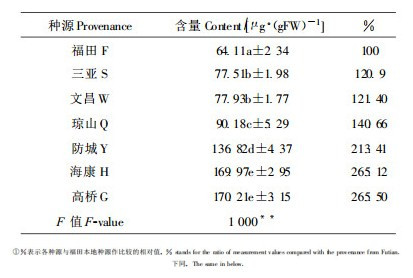
3.3 叶绿素含量叶绿素a、b分别是高等植物光合的主要色素, 其含量在一定程度上体现了光合能力大小(郑文教等, 1992)。表 4显示, 除文昌之外的种源幼苗叶绿素a、b及总量均显著高于深圳本地种源, 其中海康种源的叶绿素总量最高, 超过本地种源68.5%;方差分析和多重比较表明, 各种源间叶绿素a、b及叶绿素总量含量存在极显著差异。
|
|
林鹏等(1984)对红树植物秋茄和桐花树的叶绿素含量与土壤盐度关系研究后, 认为秋茄在盐度为0.92%时, 叶片叶绿素含量最高, 当盐度大于1.0%时, 秋茄的叶绿素合成和累积受到抑制; 污水浓度也明显影响秋茄幼苗生长和生理指标(陈桂珠等, 1994)。因此, 表 4数据很可能反映了各种源在深圳湾的特定盐度及水土污染的生境中合成和累积叶绿素量的差异, 以及对深圳湾生境的适应性之不同。
3.4 叶片过氧化氢酶活性过氧化氢酶普遍存在植物组织中, 它能催化对植物体有毒害作用的过氧化氢的分解, 增强植物的抗性(庞士铨, 1990)。不同木榄种源的一龄幼苗叶片过氧化氢酶活性为:福田 > 海康 > 高桥 > 防城 > 文昌 > 琼山 > 三亚, 各种源间的差异性达极显著水平(表 5)。这表明幼苗对体内的过氧化氢的解毒能力以福田本地种源的最强, 海康、高桥、防城种源次之, 三亚种源最弱, 这可能与福田种源长期生活在污染较严重的深圳湾而适应了这里的生境有关。
|
|
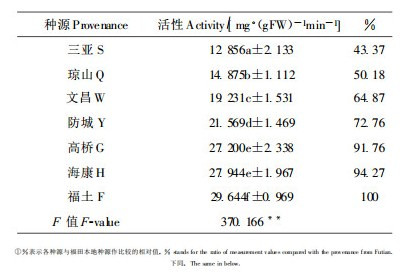
生活在干旱、盐碱环境中的植物, 体内常常积累游离脯氨酸, 其累积量与环境干旱程度、盐度和植物对干旱等的抗逆性相关(陈桂珠等, 1994)。6种外来木榄种源的幼苗叶片积累游离脯氨酸的能力明显高于深圳本地种源, 其大小为:高桥 > 海康 > 防城 > 琼山 > 文昌 > 三亚 > 福田(表 6)。这反映了各种源对生境盐度的适应能力差异。
|
|
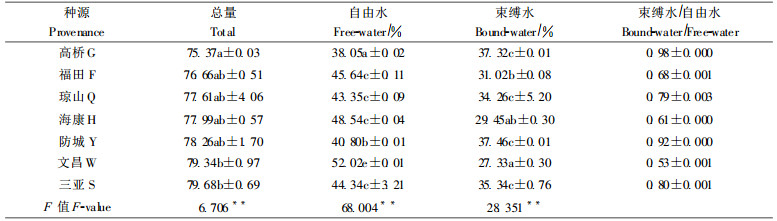
植物体内的水, 据与原生质的结合状况不同而分为自由水和束缚水。自由水含量与植物的生理活性强度有关, 它制约着光合速率、呼吸速率和生长速率; 束缚水不参与植物的代谢作用, 但与植物对不良环境的抗性有关, 当植物生长过程遇到干旱或高浓度盐水, 植物体内含水量减少, 束缚水多, 则保水力较强。在某种程度上, 体内自由水和束缚水含量及其比值与植物生长和抗性密切相关。因此, 自由水和束缚水含量是抗性生理的一个重要指标(华东师范大学生物系, 1980)。方差分析和F值检验表明, 不同木榄种源在深圳福田培育后, 幼苗含水量、自由水与束缚水比值存在极显著差异(表 7)。其中防城、三亚、琼山、高桥种源的束缚水含量明显高于其它种源, 抗逆性较强; 文昌和海康种源的幼苗束缚水/自由水值较深圳本地种源低, 而其它种源则较高。这说明文昌和海康种源在污染较严重的深圳湾生长, 其幼苗保水力较弱, 抗性较差; 而其它种源幼苗保水性较强, 抗性好。
|
|
叶片组织相对电导率是植物环境胁迫抗性指标之一(王焕校, 1990)。表 8显示, 三亚木榄种源的相对电导率最大, 海康最低; 6个外来种源中, 三亚、文昌的相对电导率高于福田本地种源, 而防城、海康、琼山、高桥种源的相对电导率低于福田本地种源; 多重比较表明各种源之间的抗逆性存在显著差异。这说明各种源幼苗对深圳湾环境胁迫的抗性存在差异。
|
|

木榄种源引种到深圳福田, 一龄幼苗在同一时刻的光合作用、呼吸作用及气孔导度有明显的不同(见表 9), 同一时刻各木榄种源幼苗光合速率以琼山、三亚较大, 高桥最小; 气孔导度以琼山、三亚、文昌、防城较大, 高桥最小; 同一时刻各种源的蒸腾速率差别较大。
|
|
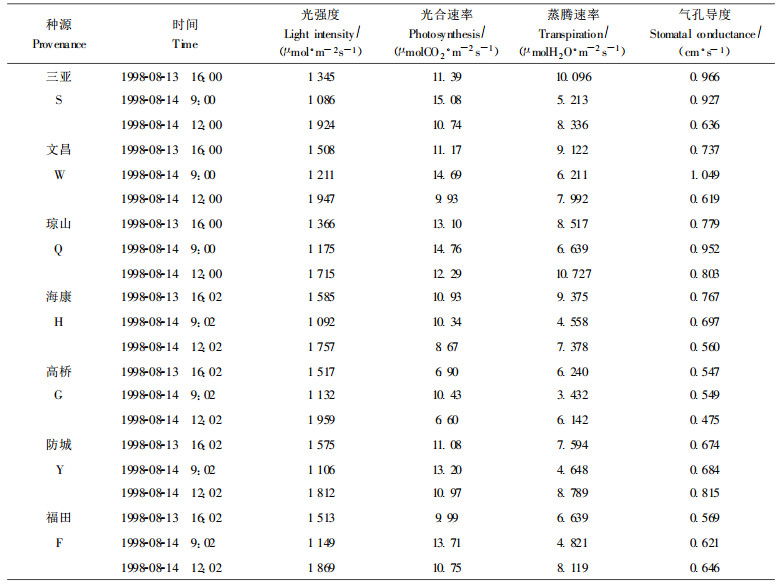
与其它陆生植物相比, 红树植物光合和蒸腾作用与土壤盐渍条件、pH值、水分条件、小气候等紧密联系(Scholander et al., 1962; 1965;Clough et al., 1992)。不同木榄种源长期适应各自生境, 引种到新的生境后, 其光合和蒸腾作用产生了一定的差异性, 这反映了各木榄种源对深圳湾生境的适应性之不同。
4 综合评定与建议综合评定是应用多维空间(欧几米德) En多向量的理论综合评定的数学模型(顾万春, 1986)。前苏联学者维列辛做欧洲赤松生态型地理造林时, 为选择优良品质的种子, 将15个生态型种子, 从6个性状方面进行综合评定, 最后确定了3个最好的种子, 该方法在前苏联应用较多(维列辛, 1978)。
不同木榄种源引种到深圳福田相同生境中, 其一龄幼苗形态、生长和生理生态指标存在不同程度的差异性, 将主要指标进行坐标综合评定, 以判别各种源品质的优劣。具体做法为:将观测值列成原始数据表, 以aij表示(i为1、2、3…7, 即代表 7个种源; j为1、2、3…9, 即代表存活率、高度、地径、叶绿素含量等9个指标), 以每列中数值最大者作比较, 得到相应位置上的相对值, 结果成为aij矩阵坐标, 用公式Pi2=∑Kij (1-aij) 2计算综合值Pi大小, 值小者为优。式中, Kij为加权系数, 考虑到各种源木榄引种到深圳福田后, 幼苗形态、生理、生态指标对其生长都比较重要, 因此, 9个指标的权重系数都定为1, 计算结果见表 10。
|
|
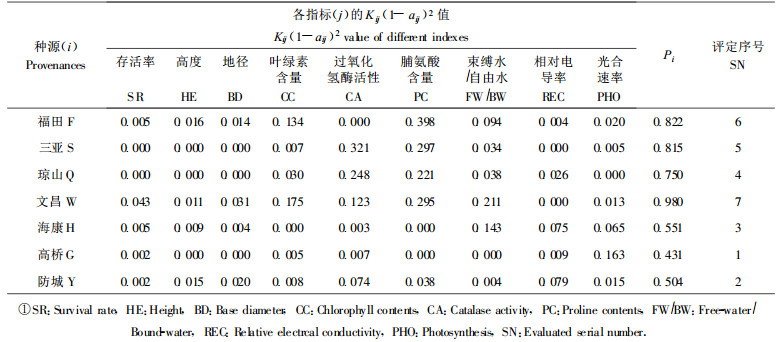
表 10中评定序号显示, 各种源综合值由低至高为:高桥、防城、海康、琼山、三亚、福田、文昌。据此确定, 高桥的木榄可能是深圳福田引种的最优种源。不过, 本研究仅仅根据种源幼苗生长、生理生态方面的实验得出初步结论, 建议继续作栽培实验和专门的种源抗逆性实验, 以更准确地判断种源的优劣。
根据我们十几年的红树林造林工作情况, 一龄幼苗是人工种植的最佳苗木, 因此, 对一龄幼苗种源的综合评定对人工造林时种源的选择有十分重要的意义。根据本文的研究结果, 建议深圳湾及周边地区种植木榄人工林时, 最好选择高桥的种源, 以促进红树林的自然恢复。
陈桂珠, 缪绅裕, 谭凤仪, 等. 1994. 人工合成污水对秋茄幼苗几个生态生理学指标影响初报. 应用生态学报, 5(2): 221-224. DOI:10.3321/j.issn:1001-9332.1994.02.022 |
顾万春. 1986. 林业试验统计. 杭州: 浙江省林业出版社, 349-353.
|
华东师范大学生物系. 1980. 植物生理学实验指导. 北京: 人民教育出版社, 5-8.
|
黄学林, 陈润政. 1990. 种子生理实验手册. 北京: 农业出版社, 122-124.
|
廖宝文, 郑德璋, 郑松发, 等. 1996. 红树植物秋茄造林技术的研究. 林业科学研究, 29(5): 575-579. |
林鹏, 陈德海, 李钨金. 1984. 两种红叶的几种酶的生理特性和海滩盐度的相关性初探. 植物生态学与地植物学丛刊, 8(3): 222-227. |
庞士铨. 1990. 植物逆境生理学基础. 哈尔滨: 东北林业大学出版社, 106.
|
王焕校. 1990. 污染生态学基础. 昆明: 云南大学出版社, 91-94.
|
王伯荪, 余世孝, 李鸣光, 等. 1996. 植物群落学实验手册. 广州: 广东高等教育出版社, 38-39.
|
维列辛(前苏联). 1978. 欧洲赤松种子评价. 森林业(前苏联), 12(1): 30-32. |
郑德璋, 郑松发, 廖宝文. 1995. 红树林湿地的利用及其保护与造林. 林业科学研究, 8(3): 322-328. |
郑文教, 林鹏. 1992. 盐度对红树植物海莲幼苗的生长和某些生理生态特性的影响. 应用生态学报, 3(1): 9-14. DOI:10.3321/j.issn:1001-9332.1992.01.014 |
朱广廉, 钟海文, 张爱琴, 等. 1990. 植物生理学实验. 北京: 北京大学出版社, 51-254.
|
朱广廉, 邓兴旺, 左卫能, 等. 1983. 植物体内游离脯氨酸的测定. 植物生理学通讯, 1(1): 35-37. |
Clough B F. Coastal and Esturine studies. In: Robertson A L, Alongi K M(eds). Tropical Mangrove Ecosystems. Washington: American Geophysical Union, DC, 1992, 225~ 250
|
Scholander P F, Hammel H T, Hemmingsen E A, et al. 1962. Salt balance in mangroves. Plant Physiol, 37: 722-729. DOI:10.1104/pp.37.6.722 |
Scholander P F, Hammel H T, Bradstreet E D, et al. 1965. Sap pressure in vascular Plant. Science, 148: 339-346. DOI:10.1126/science.148.3668.339 |