 2002, Vol. 38
2002, Vol. 38文章信息
- 何维明, 张新时.
- He Weiming, Zhang Xinshi.
- 沙地柏对毛乌素沙地3种生境中养分资源的反应
- RESPONESE OF SABINA VULGARIS TO NUTRIENT RESOURCES IN THE CONTRASTING HABITATS IN THE MU US SANDLAND
- 林业科学, 2002, 38(5): 1-6.
- Scientia Silvae Sinicae, 2002, 38(5): 1-6.
-
文章历史
- 收稿日期:2000-11-20
-
作者相关文章


2. 中国科学院植物研究所 北京 100093
2. Institute of Botany, the Chinese Academy of Sciences Beijing 100093
养分贫瘠、水分短缺、蒸散强烈是毛乌素沙地的基本环境特征。但它依然拥有非常丰富的灌木资源(张新时, 1994)。种类繁多的沙生灌木如何适应恶劣的沙地环境一直成为研究者关心的一个问题。
土壤养分状况是影响植物生长、繁殖, 生态系统净生产力等的重要因素(Larcher, 1995)。养分贫瘠是自然界普遍存在的现象。植物为适应养分贫瘠的环境常常形成许多适应性特征。例如, 常绿树种对土壤养分的利用效率高于落叶树种, 常绿树种的养分存留时间较长、养分重吸收效率较高、叶氮含量较高、叶片寿命较长等(Aerts, 1996; Santa Regina et al., 1997; Hevia et al., 1999; Eamus, 1999)。
固定沙地、流动沙地和滩地是毛乌素沙地的3种重要景观类型(陈仲新等, 1996), 三者的土壤水分含量和养分状况都存在差异(肖洪浪等, 1996)。沙地柏(Sabina vulgaris Ant.)是毛乌素沙地的天然常绿灌木(张新时, 1994), 对半干旱沙地环境具有较强的适应性。例如, 沙地柏根面积指数随土壤水分可利用性的降低而减少, 根系通过增加深度以补偿土壤水分可利用性的降低(何维明, 2000b)。本文基于野外取样和室内分析回答3个问题:(1)养分利用效率和重吸收效率是否随土壤养分可利用性的降低而提高; (2)地上部分和地下部分对养分内循环的贡献; (3)根分布与土壤养分异质性的关系。为了回答第1个问题, 作者分析了不同生境中养分利用效率和重吸收效率与土壤养分含量的关系。根和叶是植物与环境进行物质交换的窗口, 通过分析根和叶生物量和养分分配格局解决第2个问题。根据各种养分含量的变异性, 探讨根分布对土壤养分异质性的影响。
1 材料和方法 1.1 研究物种和地点沙地柏系柏科(Cupressaceae)常绿灌木, 是一种具有代表性的木本克隆植物(何维明, 2000a)。主要分布于温带大陆性干旱、半干旱区, 在中国天然分布于青海、新疆、甘肃、宁夏、内蒙古等地海拔1 100 ~ 2 800 m地区(中国植物志编委会, 1978)。研究地点位于内蒙古毛乌素沙地腹地乌审旗图克苏木(109°2′ ~ 109°17′E, 38°57′~ 39°1′N, 1 200 ~ 1 350 m)。该苏木年均降水360 mm, 80 %的降水集中于6月中旬~ 9月中旬, 年均蒸发量2 300 mm, 湿润度0.3;年均气温6.4 ℃, 1月份平均气温-11.4 ℃, 7月份平均气温21.4 ℃(姚洪林等, 1992)。
1.2 野外取样和养分分析1998-08, 分别在固定沙地(fixed sandland, FS)、流动沙地(shifting sandland, SS)和滩地(lowland, LL)上选择大小相近的沙地柏, 从基部伐倒植株, 将地上部分分成叶和茎, 将根分成粗根(直径≥2 mm)和细根(直径<2 mm)。在3种生境的沙地柏根系周围选择代表性取土点14个, 用土样盒从地表向下每隔15 cm取土样1次; 同时分别在3种生境中无沙地柏的林窗内取10个土样以作对照(垂直取样)。此外, 在固定沙地上选择具有1 m以上匍匐茎的沙地柏, 以基茎为中心沿匍匐茎每隔20 cm取土样1次, 并用相同方法在无沙地柏的林窗中水平取土20次以作对照。将地上部分放入85 ℃干燥箱, 根样品放入105 ℃干燥箱烘干, 土样在室温下风干。用电子天平(精度为0.001 g)称叶、茎、粗根和细根干重。养分含量的测定方法分别为:用重铬酸钾氧化-外加热法测土壤有机碳; 凯氏定氮法测植物和土样氮含量; 电压耦合等离子体发射光谱仪测植物和土样磷和钾含量; 原子吸收光谱法测植物和土样的钙和镁含量。养分含量测定的详细操作参见刘光崧等(1996)。计算以下各项指标:
叶质量比(leaf mass rato, LMR)=叶质量植株质量;
根(总根或细根)质量比(root mass ratio, RMR)=根(总根或细根)质量植株质量;
叶养分比(leaf nutrient ratio, LNR)=叶养分含量植株养分总量;
根(总根或细根)养分比(root nutrient ratio, RNR)=根养分含量植株养分总量;
养分利用效率(nutrient use efficiency, NUE)=凋落物量养分浓度(Vitousek, 1984);
养分重吸收效率(nutrient resorption efficiency, NRE)=(绿叶养分含量-衰老叶养分含量)绿叶养分含量(Aerts, 1997);
养分重吸收水平(nutrient resorption proficiency, NRP)=衰老叶中养分含量(Aerts, 1997)。
1.3 数据分析本文涉及3种生境, 用一元方差分析确定土壤养分含量、生物量分配、养分分配随生境的变化是否显著。然后用Duncan多重比较法确定3种生境中相应指标差异的显著性(高惠璇等, 1997)。
2 结果 2.1 土壤养分状况就0 ~ 90 cm土层养分总含量(表 1)而言, 固定沙地的氮、磷、钙含量显著高于流动沙地和滩地, 前者的钾含量显著低于后两者, 碳、镁、碳氮比随生境的变化均表现出明显的梯度性。这些结果表明, 3种生境中的土壤养分资源存在明显差异, 不同养分随生境的变化格局不相同。
|
|
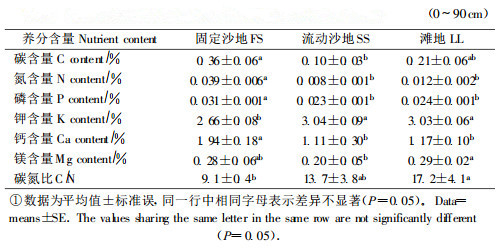
固定沙地上沙地柏叶、根和细根质量比与滩地上的沙地柏均无显著差异(图 1A、B、C)。固定沙地和滩地上的叶质量比显著小于流动沙地(图 1A), 而前两者的根质量比和细根质量比都显著大于后者(图 1B、C)。由此推断, 固定沙地和滩地上沙地柏植株生物量分配相似, 而两者与流动沙地上沙地柏生物量分配存在明显差异。

|
图 1 叶质量比、根质量比和细根质量比随生境的变化 Fig. 1 Changes in leaf mass ratio(LMR), root mass ratio(RMR) and fine root mass ratio (FRMR) with habitats.Fixed sandland(FS); shifting sandland(SS); lowland (LL). 数据为平均值±标准误, 相同字母表示差异不显著(P=0.05)。 Data=means±SE.The bars with same letter are not significantly different(P=0.05) |
表 2揭示氮、磷、钾、钙和镁在3种生境上沙地柏叶、根和细根中的分配。流动沙地上沙地柏的叶氮比、叶磷比和叶钾比分别显著大于固定沙地和滩地上沙地柏的相应值, 而这3种养分在根和细根中的分配格局恰好与叶相反; 氮、磷和钾在沙地柏叶、根中的分配随生境的变化完全相似; 根钙比不随生境发生显著变化, 但叶钙比和细根钙比随生境发生显著变化; 流动沙地上沙地柏的叶镁比显著大于固定沙地和滩地, 但根镁比和细根镁比显著小于后两者。
|
|
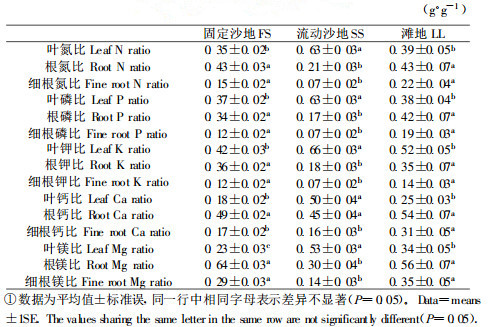
流动沙地上沙地柏植株的氮和磷利用效率分别小于固定沙地和滩地, 而前者的氮和磷重吸收效率却大于后两者; 固定沙地与流动沙地的氮、磷利用效率和重吸收效率相差最大(图 2A & B)。就钾利用效率和重吸收效率而言, 固定沙地>流动沙地>滩地(图 2C)。流动沙地上沙地柏植株的钙和镁利用效率均大于固定沙地和滩地; 固定沙地的钙利用效率高于滩地, 而镁利用效率低于滩地(图 2D & E)。

|
图 2 不同生境中的养分利用效率和重吸收效率 Fig. 2 Nutrient use efficiency and resorption efficiency under the different habitats.Fixed sandland(FS); shifting sandland(SS); lowldand(LL).  氮利用效率NUE=N use efficiency, 氮重吸收效率NRE=N resorption efficiency, 磷利用效率PUE=P use efficiency, 磷重吸收效率PRE=P resorption efficiency, 钾利用效率KUE=K use efficiency, 钾重吸收效率KRE=K resorption efficiency, 钙利用效率CaUE=Ca use efficiency, 镁利用效率MgUE=Mg use efficiency. |
本文采用养分含量变异系数反映土壤养分异质性。图 3A、B、C分别表示固定沙地、流动沙地和滩地中养分在垂直剖面上的变异性, 图 3D表示固定沙地中养分在水平剖面上的变异性。就有根系分布土壤与对照土壤而言, 无论是在垂直剖面还是水平剖面上, 养分变异系数均有不同程度的差异, 其中氮的变异系数变化最大; 垂直剖面上有根土壤的氮变异系数大于对照, 而水平剖面上恰好相反(图 3A、B、C、D)。这些结果指示, 根系分布可增大土壤氮的垂直异质性; 而减小水平异质性; 根分布可不同程度增大碳异质性, 对磷的影响较小(水平剖面除外); 根分布对钾、钙、镁的影响随生境和土壤剖面发生变化。

|
图 3 养分含量的变异系数 Fig. 3 Variance coefficients of nutrient contents □有根土壤SWR=soil with roots; 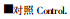
|
资源分配是一个消耗资源的过程, 分配对策体现了植物对不同环境压力作出的响应。由于生物量与碳元素存在极显著的相关, 因此生物量分配可在很大程度上反映碳元素在有机体中的分配格局(Snow et al., 1989)。生物量分配的显著差异(图 1)表明, 固定沙地和滩地上沙地柏植株的碳分配无显著差异, 而两者与流动沙地中沙地柏植株的碳分配存在显著差异。流动沙地上沙地柏的根质量比和细根质量比都小于固定沙地和滩地(图 1), 这可能与流动沙地的碳、氮和磷含量最低有关(表 1), 因为这些元素容易发生亏缺, 沙地柏植株降低根系的生物量分配有利于它更加经济、长期地利用有限的养分资源。
3.2 养分利用效率和重吸收效率与土壤养分可利用性养分利用效率常作为多年生植物对不同养分可利用性的适应指标(Aerts, 1995)。养分重吸收过程可以缓冲土壤养分波动对植物造成的伤害, 提高植物竞争力和生产力(Killingbeck, 1996; Santa Regina et al., 1997)。就土壤氮和磷含量而言, 固定沙地>滩地>流动沙地, 沙地柏叶片对氮和磷的利用效率也为固定沙地>滩地>流动沙地, 而氮和磷的重吸收效率恰好相反(表 1和图 2A, B)。这些结果指示:沙地柏正是通过提高氮和磷重吸收效率而不是通过提高氮和磷利用效率来适应氮和磷贫瘠的沙地环境的。这种现象在其它研究中已被发现(Aerts et al., 1993; Berendse, 1994)。流动沙地和滩地的钾含量高于固定沙地, 前两者叶片对钾的利用效率和重吸收效率大于后者(表 1和图 2C)。流动沙地的钙和镁含量最低, 而叶片对两者的利用效率却最高(表 1和图 4D, E)。由于钙和镁属于难再利用元素(Santa Regina et al., 1997), 沙地柏叶片对两者的适应对策是提高利用效率。钾是离子形态存在的、典型的可再利用性元素(Santa Regina et al., 1997), 沙地柏对它的适应则是提高利用效率和重吸收效率。综上所述, 沙地柏的养分利用效率和重吸收效率与土壤养分可利用性密切相关, 但对不同养分的适应性反应存在差异。

|
图 4 3种生境中的养分重吸收水平 Fig. 4 Nutrient resorption proficiency of the S. vulgaris plants in the three habitats a固定沙地, FS; b流动沙地, SS; c滩地, LL. |
在很大程度上, 多年生植物的矿质营养依赖于养分内循环(Aerts, 1997; Whigham et al., 1999)。制约养分内循环的环节很多。例如, 凋落物数量(养分归还量)、分解速率、根系吸收速率、养分在植物体的存留时间等。由于叶和根是凋落物的主要成分, 因而两者对凋落物数量和养分内循环具有重要贡献。流动沙地上沙地柏植株的叶质量比大于固定沙地和滩地(图 1), 而且前者叶片的氮、磷、钾、钙、镁比例都高于后两者(表 2), 因此流动沙地中沙地百植株叶片对养分归还的潜在贡献大于固定沙地和滩地。尽管细根生物量比例较小, 但细根周转期短, 植株同化的相当部分碳用于细根产生, 而且在很多森林中细根周转归还土壤的碳和养分数量等于或大于叶, 因此细根对养分内循环具有非常重要的作用(Hendrick et al., 1993; Jackson et al., 1997)。流动沙地中沙地柏的细根质量比和细根的氮、磷、钾、钙和镁比例都低于固定沙地和滩地(表 2和图 1)。这意味着前者细根对养分归还的潜在贡献小于后两者。
在落叶过程中, 可再利用养分有两种途径, 一部分被重吸收, 返回非凋落部分; 另一部分随凋落物归还土壤, 被分解后释放(Aerts, 1997)。凋落物中养分状况与母株养分含量和重吸收过程有关。养分重吸收水平(nutrient resorption proficiency)是指衰老叶片中养分的最终浓度(Killingbeck, 1996)。流动沙地上沙地柏植株的氮重吸收水平大于固定沙地和滩地, 磷和镁的重吸收水平非常接近, 钾的重吸收水平依次为滩地>流动沙地>固定沙地(图 4)。就叶碳氮比而言, 植株:固定沙地>滩地>流动沙地; 细根:流动沙地>固定沙地>滩地。碳氮比的提高导致多酚类碳水化合物(如纤维素、木质素)含量提高, 从而降低叶或根的分解速率(Whigham et al., 1999)。叶分解速率通常与叶氮含量成正相关(Aerts, 1997)。因此流动沙地中沙地柏植株地上部分凋落物的分解速率和养分释放量快于固定沙地和滩地, 而地下部分却恰好相反。养分重吸收和分解速率之间存在一种权衡(trade-off), 因为增大重吸收效率可提高养分利用效率, 但降低了凋落物分解速率, 不利于养分内循环速率的提高(Aerts, 1997)。由于流动沙地上沙地柏植株叶片对养分归还数量的潜在贡献大于固定沙地和滩地, 而且前者的分解速率也快于后两者, 因此流动沙地中植株叶片对养分内循环的贡献大于固定沙地和滩地。然而, 流动沙地上植株根系对养分内循环的贡献却小于固定沙地和滩地, 因为前者的养分归还量较少、而且分解速率也较低。这暗示沙地柏殖株叶片和根系对养分内循环的贡献可能存在一种权衡, 即如何分配叶和根对养分内循环的贡献。
3.4 根分布与土壤养分异质性土壤是异质性环境。产生养分异质性的原因多种多样, 如养分归还速率、凋落物分解、根系选择性吸收、微地形、干扰等(Bazzaz, 1996; Ryel et al., 1998)。在垂直剖面上, 沙地柏根系分布增大了土壤碳、氮、钙和镁的异质性程度(滩地上的钙、镁相反), 对磷异质性的影响很小, 使钾的同质性增强; 在水平剖面上, 根系分布效应的总趋势是降低土壤养分异质性(图 3)。由此可以初步推断, 沙地柏根系分布可改变土壤养分的分布状况, 根系分布对土壤养分水平异质性和垂直异质性的影响也存在差异。根系分布与土壤养分的关系非常复杂, 因为养分状况影响根系分布, 而根系分布反过来影响土壤养分可利用性。开展这方面的定量研究有助于更好地理解植物根系分布与养分异质性之间的关系。
陈仲新, 张新时. 1996. 毛乌素沙地景观生态分类与排序的研究. 植物生态学报, 20: 423-437. |
高惠璇, 耿直, 李贵斌, 等. 1997. SAS系统. SAS STAT软件使用手册. 北京: 中国统计出版社, 250-268.
|
何维明. 2000a. 切断匍匐茎对沙地柏子株生长和资源利用效率的影响. 植物生态学报, 24: 391-395. |
何维明. 2000b. 不同生境中沙地柏根面积分布特征. 林业科学, 36(5): 17-21. |
刘光崧, 蒋能慧, 张连低, 等. 1996. 土壤理论分析与剖面描述. 北京: 中国标准出版社, 11-28.
|
肖洪浪, 张继贤, 李金贵. 1996. 沙漠流动沙地固定过程中土壤肥力演变. 中国沙漠, 16(增刊1): 64-69. |
姚洪林, 魏成泰, 廖茂彩. 内蒙古毛乌素沙地开发整治研究中心概况. 见: 王家祥, 姚洪林, 魏成泰. 毛乌素沙地开发整治研究中心研究论文. 呼和浩特: 内蒙古大学出版社, 1992, 1~ 8
|
张新时. 1994. 毛乌素沙地的生态背景及其草地建设原则与优化模式. 植物生态学报, 18: 1-16. DOI:10.3321/j.issn:1005-264X.1994.01.008 |
中国植物志编委会. 1978. 中国植物志(第七卷). 北京: 科学出版社, 359-360.
|
Aerts R. 1995. The advantages of being evergreen. Trends in Ecology and Evolution, 10: 502-507. |
Aerts R. 1996. Nutrient resorption from senescing leaves of perennials: are there general patterns?. Journal of Ecology, 84(597): 608. |
Aerts R. 1997. Nitrogen partitioning between resorption and docomposition pahtways: a trade-off between nitrogen use efficiency and litter decomposibility?. Oikos, 80(306): 606. |
Aerts R, van der Peijl M J. 1993. A simple model to explain the dominance of low-productivity perennials in nutrient-poor habitats. Oikos, 66: 144-147. DOI:10.2307/3545208 |
Bazzaz F A. 1996. Plants in changing environments. London: Academic Press, 28-49.
|
Berendse F. 1994. Competition between plant population at high and low nutrient supply. Oikos, 71: 253-260. DOI:10.2307/3546273 |
Eamus D. 1999. Ecophysiological traits of deciduous and evergreen woody species in the seasonally dry tropics. Trends in Ecology and Evolution, 14: 11-16. DOI:10.1016/S0169-5347(98)01532-8 |
Hendrick R L, Prgitzer K S. 1993. Patterns of fine root mortality in two sugar maple forests. Nature, 361: 59-61. DOI:10.1038/361059a0 |
Hevia F, Minoletti M L, Decker K L M , et al. 1999. Foliar nitrogen and phosphorus dynamics for three chilean Nothofagus(Fagaceae)species in relation to leaflifespan. American Journal of Botany, 86: 447-455. DOI:10.2307/2656765 |
Jackson R B, Mooney H A, Schulze E-D. 1997. A global budget for fine root biomass, surface area, and nutrient contents. Proceedings of National Science, USA, 94: 7362-7366. DOI:10.1073/pnas.94.14.7362 |
Killingbeck K T. 1996. Nutrients in senesced leaves: keys to the search for potential resorphion and resorption proficiency. Ecology, 77: 1716-1726. DOI:10.2307/2265777 |
Larcher W. 1996. Physiological Plant Ecology. Berlin: Springer-Verlag, 25-46.
|
Ryel R J, Caldwell M M. 1998. Nutrient acquisition from soils with patchy nutrient distributions as assessed simulation models. Ecology, 79: 2735-2744. DOI:10.1890/0012-9658(1998)079[2735:NAFSWP]2.0.CO;2 |
Santa Regina I, Rico M, Rapp M, et al. 1997. Seasonal variation in nutrient concentration in leaves and branches. Journal of Vegetation Science, 8: 651-654. DOI:10.2307/3237369 |
Snow A A, Whigham D F. 1989. Costs of flower and fruit production in Tipularia discolor (Orchidaceae). Ecology, 70: 1286-1293. DOI:10.2307/1938188 |
Vitousek P M. 1984. Litterfall, nutrient cycling, and nutrient use efficiency. Ecology, 65: 285-298. DOI:10.2307/1939481 |
Whigham D E, Twilley R R, Feller I C. 1999. Within-stand nutrient cycling in wetland ecosystems. Ecology, 80: 2137-2138. |