 2002, Vol. 38
2002, Vol. 38文章信息
- 吴承祯, 洪伟.
- Wu Chengzhen, Hong Wei.
- 珍稀濒危植物长苞铁杉种群密度效应模型
- A STUDY ON THE DENSITY EFFECT MODEL OF RARE AND ENDANGERED TSUGA LONGIBRACTEATA POPULATION
- 林业科学, 2002, 38(4): 157-161.
- Scientia Silvae Sinicae, 2002, 38(4): 157-161.
-
文章历史
- 收稿日期:2001-04-09
-
作者相关文章


植物种群的调节是一个多因素的复杂过程, 通过对种群的出生、死亡和生长等过程的控制, 使种群的数量维持在生境可以容纳的最大水平上。尽管影响种群动态的因素可分为密度制约因素和非密度制约因素, 但在植物中, 尤其对木本植物, 主要是密度制约起着种群调节与控制的作用, 这种调控的过程反映了种群对群落结构及相应环境条件的适应与利用, 主要表现为植物种群密度效应(江洪, 1992)。种群密度效应一直是种群生态学领域研究的热点。前人在研究植物种群密度拥挤效应规律时, 提出了大量理论或经验的表达式(Yada et al., 1963; 唐守正, 1993; 方精云, 1995; Hozumi, 1977; 1983;John et al., 1994; 张大勇等, 1985; 吴承祯等, 2000c; 2000d;安藤贵, 1968; 洪伟等, 1996; 陈辉等, 1992), 反映了植物种群数量动态受多种限制因素影响, 如有限的资源、空间以及种内、种间植物个体对资源的竞争, 导致种群数量发生变化, 即为密度效应。
美国Rieneke首先提出了林分密度竞争效应的乘幂式:
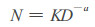
|
(1) |
其中:N为种群密度; D为平均直径; a、K为因树种而变化的常数。
日本的吉良龙夫(Yada et al., 1963)发现草木植物密度与生长之间存在:
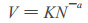
|
(2) |
其中:V为平均个体重量; N为单位面积株数。1982年Silvertoan在研究林分自然稀疏时, 也同样提出这种模型。但模型(2)必须在林分疏密度相同时使用才有意义。
安藤贵认为林分生长过程中, 不断分化出来的生长落后的被压木, 因光照和营养不足, 最后导致自然枯死(安藤贵, 1968)。于是提出:

|
(3) |
立木密度与林分单株材积产量的密度效应关系也可用密度二次效应模型表示:
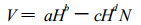
|
(4) |
前人研究中较多地应用(2)式来研究森林自疏和密度效应规律, O' Neill和DeAngelis研究娑罗双和柚木, 赵学农(1991)研究木果石栎均采用(2)式。但是由于模型(2)必须在林分疏密度相同时才有意义, 因此, 陈辉等(1992)提出密度效应新模型:
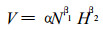
|
(5) |
#其中:V为单株平均材积; N为单位面积密度; H为林分的优势木平均高; α、β1、β2为模型参数。
笔者(吴承祯等, 2000d)曾提出一个能描述植物种群密度变化全过程的密度效应模型:
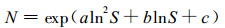
|
(6) |
其中:N为单位面积株数(株·hm-2); S为林分单株平均胸高断面积(m2·株-1); a、b、c为待定参数。
长苞铁杉(Tsuga longibracteata)属松科铁杉属, 为国家三级保护植物(国家环境保护局, 1987), 是珍贵稀有树种之一。其分布范围相对狭窄, 散生于我国的贵州、湖南、广东、广西和福建等地(林金星等, 1995)。目前长苞铁杉的残存个体数量有限, 在福建省天宝岩自然保护区保存有一片较完好的长苞铁杉针阔混交林, 面积约72 hm2左右。因此, 研究其种群生态学与群落生态学, 对于保护珍稀濒危植物长苞铁杉具有一定的生态意义和理论价值。但有关长苞铁杉种群生态学与群落生态学研究报道较少(吴承祯等, 2000a; 2000b;2001), 本文探讨长苞铁杉种群密度效应, 旨在考察长苞铁杉种群密度调节的过程与机制、揭示其在不同自疏阶段中的密度变化规律。
1 研究区概况福建省天宝岩自然保护区位于北纬25°55′~ 25°58′, 东经117°31′~ 117°33.5′, 面积约1976.5 hm2, 是戴云山余脉, 为中低山地貌, 最高峰天宝岩1604.8 m。保护区年平均温度23 ℃左右, 最冷月(1月)绝对最低温度-11 ℃, 最热月(7月)绝对最高温度40 ℃, 全年≥10 ℃的活动积温在4520 ~ 5800 ℃左右, 年降雨量2000mm, 空气相对温度80 %左右。土壤的垂直分布大致是海拔800 m以下为红壤, 800 ~ 1350 m为黄红壤, 1350 m以上为黄壤。长苞铁杉群落外貌整齐, 色彩暗绿, 林内阴暗湿润, 土层较厚, 表土质地松软, 枯枝落叶层厚3 ~ 5cm, 为中亚热带山地垂直地带性植被类型。长苞铁杉主要伴生树种有甜槠(Castanopsis eyrei)、杉木(Cunninghamia lanceolata)、木荷(Schima superba)、深山含笑(Michelia maudiae)、薯豆(Elaeocarpus japonica)、小叶青冈(Cyclobalanopsis myrsinaefolia)、冬青(Ilex purpurea)及猴头杜鹃(Rhododendron simiarum)等。
2 材料与方法 2.1 调查方法在保护区内选择生长发育正常的长苞铁杉天然混交林, 依不同生境条件、不同发育阶段设置8块样地, 每块样地面积600 m2。对样地内林木进行每木检尺, 记录样地所有个体的种名、胸径、树高、冠幅(起测径阶≥4 cm), 同时每100 m2选择1株长苞铁杉优势木计算优势木平均高; 每一样地内随机设置6个1 m×1 m小样方调查长苞铁杉群落内灌木和幼苗幼树种类、高度及盖度、分布情况。8块样地基本情况如表 1。
|
|
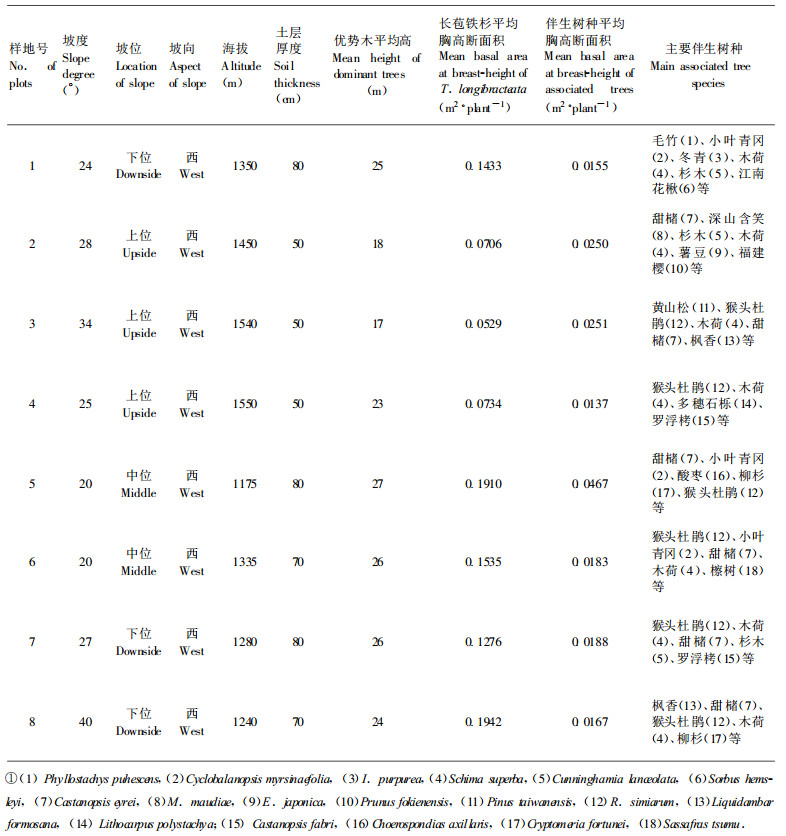
考虑到各模型本身的特性及其适应性, 本文应用模型(6)来研究珍稀濒危植物长苞铁杉种群密度效应规律, 一方面动态描述长苞铁杉种群密度调节过程与机制, 另一方面进一步检验本文所提出的密度效应模型的适应性。
3 结果与分析由(6)式可知, 此方程为非线性方程, 非线性方程一般采用麦夸方法拟合。但麦夸方法复杂, 需要求偏导, 因此计算量大而繁琐。本文采用遗传算法进行最优拟合(吴承祯等, 1997; 1998)。
根据遗传算法的基本思想, 应用BASIC语言编制了长苞铁杉种群密度效应规律模拟的遗传算法计算机运行程序, 经计算机运算得到长苞铁杉种群密度效应模型:
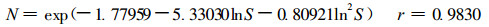
|
(7) |
式中, N为长苞铁杉种群密度(株·hm-2); S为平均单株胸高断面积(m2·株-1)。该式表明随着单位面积株数增加, 种群平均单株胸高断面积逐步减小, 较符合植物种群密度效应规律。但长苞铁杉种群生长受环境和年龄影响, 不同生境、不同年龄间长苞铁杉种群生长必然存在差异, 即使种群密度一致, 长苞铁杉种群平均胸高断面积也有差异。因此, 尽管模型(7)模型相关指数较大, 但从理论上说用(6)式研究长苞铁杉林各林分类型显然不适宜。考虑到种群密度随林分生长、立地条件的变化而变化, 引入用长苞铁杉优势木平均高H表示的不同立地条件林分类型参数值:
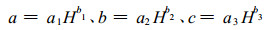
|
这样得到:

|
(8) |
模型中H集中反映林地立地质量, S可以反映不同发育阶段林木的平均大小, 因此从理论上讲该模型能客观地描述植物种群随立地、发育阶段而变化的密度效应规律。
经遗传算法最优拟合, 得到随优势木平均高变化的密度效应规律模型为:

|
(9) |
生境不同, 种群密度调节控制的强度也不同, 由于模型(9)综合考虑林分类型长苞铁杉优势木平均高, 因此较好地反映了生境立地质量对种群调控能力的影响。根据计算, 模型残差平方和为1906.27, 仅为模型(7)的6.64 %, 说明用长苞铁杉优势木平均高改进长苞铁杉种群密度拥挤效应规律模型效果较好。
由于长苞铁杉林由多树种组成, 长苞铁杉种群动态不仅受其自身的种内竞争的影响, 还受伴生树种对其种间关系的影响, 为此, 本研究进一步探讨长苞铁杉种群伴生树种对其密度效应规律的影响机制, 即在其密度效应规律模型中增加伴生树种为影响因子。研究表明伴生树种平均单株胸高断面积对长苞铁杉种群密度存在显著的影响, 而伴生树种优势木平均高对其影响不显著。经遗传算法拟合得到考虑其伴生树种影响的长苞铁杉种群密度效应规律模型:
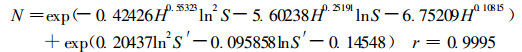
|
(10) |
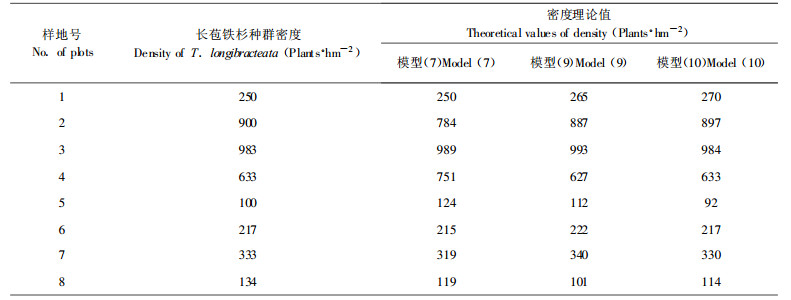
其中:S′为伴生树种平均单株胸高断面积(m2·株-1)。模型(10)的残差平方和为930.21, 仅为模型(7)的3.24 %、模型(9)的48.80 %; 利用模型(7)、(9)、(10)计算长苞铁杉种群密度理论值(表 2), 表明模型(10)的效果最好。可知在探讨长苞铁杉种群密度效应时, 考虑伴生树种对其影响是必要的。
|
|
密度效应反映由各种密度所形成的群体以及组成群体的个体之间相互作用的规律。长苞铁杉群落为多树种组成的异龄林, 每个个体在群落内的地位和作用不同, 对资源环境的要求也不相同。在相同资源条件下, 生活力强、遗传性强的个体尽可能占据林冠层, 获取尽可能多的资源, 并控制其它个体对资源的获取, 成为支配者; 生长弱小、遗传性差的个体, 因竞争能力远不及较大个体, 只能占据剩余空间和依靠剩余资源生存, 处于林冠下层, 生长发育受到抑制, 从而被淘汰。而处于林冠层上层的长苞铁杉个体, 由于生长发育旺盛, 占据了大部分营养空间; 加之与伴生树种间的种间竞争, 导致处于劣势的长苞铁杉种群个体生长受抑并逐渐被淘汰乃至死亡, 从而使得林分中长苞铁杉种密度下降, 其平均胸高断面积则随之上升。利用收集的长苞铁杉种群密度变化资料建立了其密度效应规律模型, 并考虑用长苞铁杉优势木平均高和伴生树种胸高断面积指标改进其密度效应模型, 改进后的模型相关指数为0.9995, 残差平方和仅为原模型的3.24 %, 拟合效果较好, 模型较好地描述了长苞铁杉种群密度效应规律, 可应用于指导其密度调节与控制。改进模型模拟表明, 长苞铁杉种群密度变化机制既受其自疏作用的影响, 也受其伴生树种他疏作用的制约。由于模型(6)中的参数a、b、c反映了植物种群密度效应受树种特性影响的制约程度, 而根据所建立的改进模型(10)式, 不难发现长苞铁杉种群所对应的模型制约参数值的负值程度均比其伴生树种对应的模型制约参数值大, 因此可以认为长苞铁杉种群密度效应受其自身的制约作用要比受其伴生树种的制约大得多。由此, 我们有理由认为长苞铁杉种群密度变化过程中的自疏作用大于其伴生树种的他疏作用, 自疏是影响长苞铁杉种群密度减少的主要原因。
本文建立的密度效应规律模型(10)式由于既考虑了立地质量(平均优势高)、又考虑了种群平均大小(平均胸高断面积), 因此适用于不同立地条件、不同类型林分的各发育阶段的长苞铁杉种群密度效应规律研究; 不仅克服了前人模型只适用于某一林分整个生长过程的密度效应规律研究的局限性, 而且充分考虑了种间竞争对密度效应规律的影响。但是由于该模型为非线性模型, 因此必须选择适当的非线性拟合方法加以拟合, 非线性模型的拟合方法有夸麦方法、三次设计法、改进单纯形法等方法, 本文提出采用遗传算法实现模型的最优拟合。实例拟合结果表明, 该方法具有较强的非线性拟合能力, 是生态学、生物学及林学领域大量非线性模型的理想参数估计方法。
安藤贵. 1968. 同龄单纯林の密度管理に关すゐ生态学的研究. 林试研报, 210: 1-152. |
陈辉, 何宗明, 洪伟. 1992. 杉木人工林密度效应模型的研究. 福建林学院学报, 12(3): 277-282. |
方精云. 1995. 一种描述植物种群自然稀疏过程的经验模型. 林业科学, 31(3): 247-253. |
国家环境保护局, 中国科学院植物研究所. 1987. 中国珍稀濒危植物名录. 第一册. 北京: 科学出版社, 68.
|
洪伟, 吴承祯. 1996. 闽北杉木人工林密度效应新模型. 浙江林学院学报, 13(1): 15-20. |
林金星, 胡玉熹, 王献溥, 等. 1995. 中国特有植物长苞铁杉的生物学特性及其保护. 生物多样性, 3(3): 147-152. |
江洪. 1992. 云杉种群生态学. 北京: 中国林业出版社, 51-92.
|
唐守正. 1993. 同龄纯林自然稀疏规律的研究. 林业科学, 29(3): 234-241. |
吴承祯, 洪伟. 1997. 用遗传算法改进约束条件下造林规划设计研究. 林业科学, 34(2): 133-141. |
吴承祯, 洪伟. 1998. 人工林经营过程密度最优控制研究. 自然资源学报, 13(2): 175-180. |
吴承祯, 洪伟, 吴继林, 等. 2000a. 珍稀濒危植物长苞铁杉种群生命表分析. 应用生态学报, 11(3): 333-336. |
吴承祯, 洪伟, 吴继林, 等. 2000b. 珍稀濒危植物长苞铁杉的分布格局. 植物资源与环境学报, 9(1): 31-34. |
吴承祯, 洪伟. 2000c. 杉木林自疏过程密度调节规律的研究. 热带亚热带植物学报, 8(1): 28-34. |
吴承祯, 洪伟. 2000d. 杉木人工林自疏规律研究. 林业科学, 36(4): 97-101. |
吴承祯, 洪伟, 吴继林, 等. 2001. 长苞铁杉群落种间竞争的研究. 西北植物学报, 21(1): 154-158. |
赵学农. 1991. 哀牢山果石栎林种群调节与竞争的初步研究. 植物生态学与地植物学学报, 15(2): 183-189. |
张大勇, 赵松龄. 1985. 森林自疏过程中密度变化规律的研究. 林业科学, 21(4): 369-374. |
Hozumi K. 1977. Ecological and mathematical considerations on self-thinning in ever-aged pure standsⅠ, Mean plant weight-density trajectory during the course of self-thinning. Bot.Mag.Tokyo, 90: 165-179. DOI:10.1007/BF02488355 |
Hozumi K. 1983. Ecological and mathematical considerations on self-thinning in ever-aged pure stands Ⅲ, Effect of the linear growth factor on self-thinning. Bot.Mag.Tokyo, 96: 171-191. DOI:10.1007/BF02498999 |
John P M, William F S. 1994. Stand and tree dynamics of uneven-aged Ponderosa pines For. Sci., 40(2): 189-302. |
Yada K, Kira T, Ogawa H, et al. 1963. Self-thinning in overcrowded pure stands under cultivated and natural conditions. J.Biol.Osaya.City Univ., 14: 107-129. |