 2002, Vol. 38
2002, Vol. 38文章信息
- 尹淑艳, 孙绪艮.
- Yin Shuyan, Sun Xugen.
- 针叶小爪螨—寄主植物—芬兰钝绥螨相互关系的研究Ⅲ.寄主植物的化学组成与针叶小爪螨生长发育的关系
- STUDIES ON THE INTERRELATIONSHIP AMONG OLIGONYCHUS UNUNGUIS, HOST PLANTS AND AMBLYSEIUS FINLANDICUS Ⅲ.RELATIONSHIP BETWEEN CHEMICAL COMPONENT OF HOST PLANTS AND DEVELOPMENT OF OLIGONYCHUS UNUNGUIS
- 林业科学, 2002, 38(4): 105-110.
- Scientia Silvae Sinicae, 2002, 38(4): 105-110.
-
文章历史
- 收稿日期:2000-09-11
-
作者相关文章



2. 山东农业大学林学院 泰安 271018
2. College of Forestry, Shandong Agricultural University Taian 271018
植物对植食性昆虫的防御包括化学防御和形态构造防御, 化学防御机制的研究是当前迅速发展的化学生态学中的一个重要部分(钦俊德, 1987)。化学抗性可包括两个方面, 即组成抗性和诱导抗性。组成抗性是指植物在遭到植食性昆虫进攻前就已存在的抗虫特性; 诱导抗性是指植物在遭受植食性昆虫进攻后所表现出来的一种抗虫特性。诱导抗性又可分为直接的和间接的。直接的诱导抗性是指植物受植食性昆虫危害诱导发生的变化直接对植食性昆虫产生不利影响; 间接的诱导抗性是指植物受植食性昆虫为害诱导发生的变化引诱植食性昆虫的天敌, 使植物免受更大的危害(Dicke et al., 1988)。植食性螨类的取食危害也会使植物产生诱导抗性。如:朱砂叶螨(Tetranychus cinnabarinus)取食茄子叶片, 诱导叶片内的单宁酸含量增加, 当单宁酸的含量增加到某个阈值时, 叶螨的种群数量迅速下降(王海波等, 1993);日本的Takeshi shimoda等(1997)研究证明, 菜豆受二斑叶螨(Tetranychus urticae)危害后, 释放出一些挥发性物质, 能吸引二斑叶螨的天敌—塔六点蓟马(Scolothrips takahashii)。
本文测定了不同寄主植物和同一寄主植物受害与否对针叶小爪螨生长发育的影响及相应植物叶片中几种化学物质的含量, 以揭示寄主植物对针叶小爪螨的化学防御机制。
1 材料和方法虫源 针叶小爪螨采自泰山上的麻栎(Quercus acutissima Carr.)、栓皮栎(Quercus variabilis BL.)、板栗(Castanea mollissima BL.)。
供试寄主植物叶片 采自泰山上的板栗、麻栎、栓皮栎和槲树(Quercus dentata Thunb.)。
受害叶片制备 采取未受害枝条, 置于盛有1 %蔗糖溶液的广口瓶内保鲜。在未受害叶片上接上不同螨态的针叶小爪螨, 每叶不少于50头, 危害5 d后将叶片上各发育阶段的叶螨及丝网、排泄物、卵、卵壳等用小毛笔刷除。
饲养 用海绵水盘法(孙绪艮, 1990)饲养。将叶片用干净的湿纱布擦净, 控制有效叶片的面积为4cm2, 将刚孵化的幼螨单头接到叶片上, 放于28 ℃的光照培养箱内, 每天定时(7:00, 19:00)观察两次, 记录各个螨态的发育历期、发育到成螨前的死亡率、成螨寿命、雌成螨产卵量等。每种植物叶片饲养观察叶螨50头。
叶片化学物质的测定 将待测叶片用湿纱布擦净, 剪除叶柄, 置55 ℃烘箱中烘干至恒重, 用粉碎机粉碎, 装入磨口广口瓶中待测。
游离氨基酸用氨基酸自动分析仪测定; 可溶性总糖的测定用斐林-碘量法; 氮的测定用凯氏定氮法; 粗脂肪的测定用残余法; 单宁的测定用高锰酸钾氧化法。
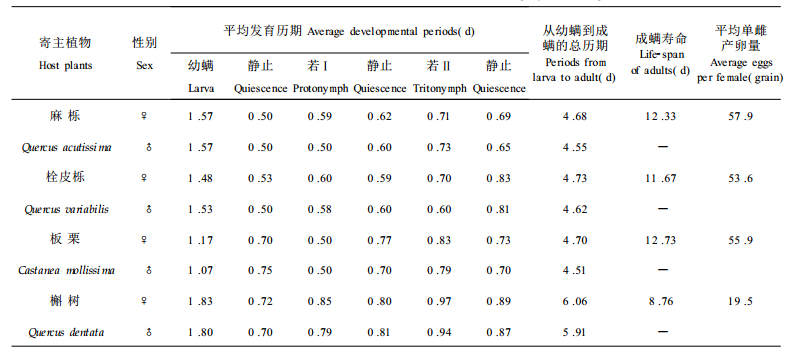
2 结果与分析 2.1 不同寄主植物对针叶小爪螨生长发育的影响用不同寄主植物的未受害叶片单头饲养针叶小爪螨, 观察各发育阶段的历期、成螨寿命、平均单雌产卵量等, 结果见表 1。由表 1可看出, 取食不同的寄主植物, 针叶小爪螨各螨态的发育历期、成螨寿命及平均单雌产卵量不同。其中取食槲树的针叶小爪螨从幼螨发育到成螨的历期最长, 雌成螨寿命最短, 平均单雌产卵量最少; 取食麻栎、栓皮栎、板栗的针叶小爪螨从幼螨发育到成螨的历期、雌成螨寿命、平均单雌产卵量之间无明显差异, 但是取食板栗的针叶小爪螨的幼螨期明显缩短。
|
|
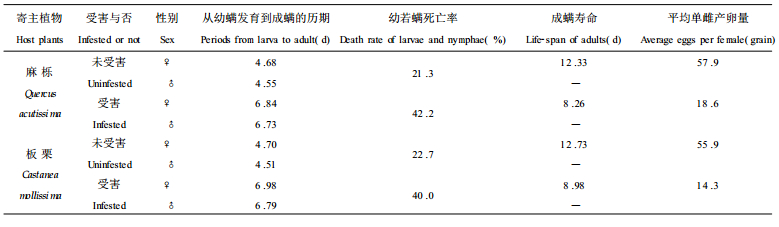
由表 2可看出, 叶片受害与否, 对针叶小爪螨的发育历期、死亡率、成螨寿命、平均单雌产卵量有明显影响。取食受害叶片的针叶小爪螨, 从幼螨发育到成螨的历期明显延长(比取食未受害叶片的针叶小爪螨延长了约2.2 d), 成螨寿命明显缩短(比取食未受害叶片缩短了约4 d), 平均单雌产卵量明显减少(约是取食未受害叶片的1/3), 死亡率明显升高(约是取食未受害叶片的2倍)。
|
|
寄主植物的营养状况及所含化学物质的种类和数量对昆虫的生长、发育、繁殖和存活有重要作用(邹运鼎等, 1992)。含氮量是限制植食性昆虫生长、发育和繁殖的关键因子(吴坤君等, 1993; Mattson et al., 1980), 害虫的发生和危害程度与寄主植物的含氮总量有关(曾益良等, 1982; Lightfoot et al., 1987), 也与寄主植物中可溶性糖的含量有密切关系(钦俊德等, 1962; 吴坤君等, 1992)。氮和糖是食物中的两类基本物质, 通过一系列的消化、吸收、分解和合成过程, 前者成为虫体的组成部分, 后者主要提供昆虫生命活动所需的能量并影响昆虫的生长发育和生殖(吴坤君等, 1992; 娄永根等, 1997)。叶组织中可溶性碳水化合物和游离氨基酸的种类和含量可决定植物对昆虫的抗性和防御能力(胡萃等, 1997)。
单宁是植物次生代谢过程中产生的化学物质, 其分子芳香环上的羟基可与蛋白质分子羧基结合形成稳定的交叉链, 所以能抑制酶的活性, 或使蛋白质鞣化。单宁不仅能与蛋白质络合, 并且能与淀粉络合而影响昆虫对淀粉的消化, 因此, 单宁是一类有效的化学防御物质(钦俊德, 1987)。
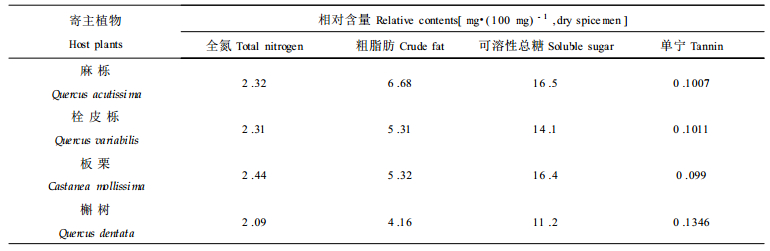
由表 3和表 4可见, 麻栎、栓皮栎、板栗叶片中游离氨基酸的总量、全氮含量和可溶性总糖的含量均较高, 能够满足针叶小爪螨生长发育的需要, 因此取食麻栎、栓皮栎、板栗的针叶小爪螨的生长发育状况均较好, 从幼螨发育到成螨的历期、雌成螨寿命、平均单雌产卵量在3者之间无明显差异。但是板栗叶片中游离氨基酸的总量和全氮含量更高些, 取食板栗的针叶小爪螨的幼螨期比取食麻栎、栓皮栎的明显缩短, 说明幼螨对植物体中的含氮量比较敏感, 含氮量高能够加速幼螨的发育。在麻栎、栓皮栎、板栗和槲树这4种寄主植物中, 槲树叶片中游离氨基酸的总量、全氮含量、可溶性总糖的含量最低, 单宁含量最高。与其他3种寄主植物相比, 取食槲树的针叶小爪螨的发育历期最长, 成螨寿命最短, 平均单雌产卵量最少, 在野外的发生量也最轻, 这可能与槲树叶片中营养物质含量低、抗虫物质含量高有直接关系。
|
|

|
|
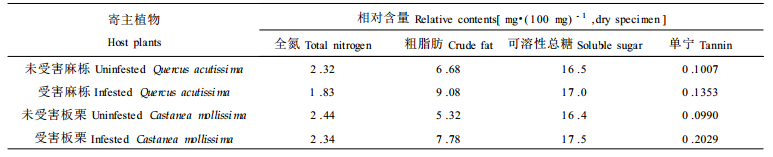
植物受到昆虫的危害后, 其体内化学物质的种类及数量会发生变化, 对昆虫产生忌避作用或使其中毒。这些变化包括营养物质含量、比例的改变以及次生代谢的加强, 诱导产生高浓度的抗虫物质(李镇宇等, 1998)。由表 5和表 6可见, 叶片受针叶小爪螨为害后, 17种游离氨基酸中, 除了蛋氨酸的含量有所增加外, 其他几种游离氨基酸的含量均减少, 游离氨基酸的总量和全氮含量减少, 可溶性总糖的含量增加, 使得氮糖比值减小, 粗脂肪和单宁的含量明显增加。受害麻栎叶片中粗脂肪的含量比未受害麻栎叶片增加了35.9 %, 单宁含量增加了34.4 %; 受害板栗叶片中粗脂肪的含量比未受害板栗叶片增加了46.2 %, 单宁含量增加了104.9 %。生物测试结果表明, 取食受害叶片与未受害叶片的针叶小爪螨相比, 发育历期延长, 成螨寿命缩短, 平均单雌产卵量减少, 死亡率升高。其原因可能是:①受害叶片营养恶化, 营养物质含量比例失调, 不利于针叶小爪螨的生长发育; ②受害叶片单宁含量明显增加, 影响了针叶小爪螨对食物的消化和利用, 从而阻碍了它们的生长发育, 降低繁殖力; ③粗脂肪包含一大类物质, 如脂肪、磷脂、酯、固醇、芳香油、色素及有机酸等, 已知芳香油及某些有机酸(癸酸、辛酸和己酸等)对昆虫是有毒的(钦俊德, 1987)。受害叶片中粗脂肪含量明显增加, 对针叶小爪螨有毒的物质含量也会随之增多, 这些物质在螨体内积累到一定程度导致叶螨生长缓慢, 甚至死亡。
|
|

|
|
野外调查中发现, 针叶小爪螨在槲树上发生较轻。室内饲养测试的结果为:取食槲树的针叶小爪螨与取食麻栎、栓皮栎、板栗的针叶小爪螨相比, 其从幼螨发育到成螨的历期最长, 雌成螨寿命最短, 平均单雌产卵量最少。这些说明槲树对针叶小爪螨有较强的抗性。在麻栎、栓皮栎、板栗、槲树4种寄主植物中, 槲树叶片中游离氨基酸的总量、全氮含量、可溶性总糖的含量最低, 单宁含量最高。几种主要的营养物质含量过低, 抗虫物质单宁含量高可能是槲树抗性形成的重要原因之一。槲树对针叶小爪螨的抗性, 除了营养物质、单宁的作用外, 其它抗虫次生代谢物质及形态结构的作用有待于研究。进一步明确槲树的抗性机理, 可望在培育抗性品种、开发天然生物活性物质防治针叶小爪螨方面提供理论依据。
针叶小爪螨的危害能使其寄主植物产生诱导抗性。表现为取食同一寄主植物受害叶片的针叶小爪螨与取食未受害叶片的相比, 其从幼螨发育到成螨的历期延长, 雌成螨寿命缩短, 平均单雌产卵量减少, 死亡率升高。
植物产生诱导抗性的原因, 是因为植物在遭受植食性昆虫的危害后, 会在各个方面包括个体发育、形态特征、生理状况、营养成分及次生化合物发生变化, 因而影响植食性昆虫和其天敌的行为(娄永根等, 1997)。板栗、麻栎受针叶小爪螨为害后, 叶片中游离氨基酸的总量、全氮含量减少, 可溶性总糖的含量增加, 氮糖比值减小, 粗脂肪和单宁的含量明显增加。
胡萃, 杨樟法. 1997. 亚洲玉米螟对氮、糖的吸收和利用. 昆虫学报, 40(2): 151-157. DOI:10.3321/j.issn:0454-6296.1997.02.006 |
李镇宇, 陈华盛, 袁小环, 等. 1998. 油松对赤松毛虫的诱导化学防御. 林业科学, 34(2): 43-49. DOI:10.3321/j.issn:1001-7488.1998.02.007 |
娄永根, 程家安. 1997. 植物的诱导抗虫性. 昆虫学报, 40(3): 320-330. DOI:10.3321/j.issn:0454-6296.1997.03.018 |
钦俊德, 李丽英, 魏定义, 等. 1962. 关于棉铃虫食性和营养的某些特点. 昆虫学报, 11(4): 327-340. |
钦俊德. 1987. 昆虫与植物的关系. 北京: 科学出版社.
|
孙绪艮. 1990. 针叶小爪螨研究初报. 山东农业大学学报, 21(3): 41-46. |
王海波, 吴千红, 高闻达. 1993. 茄子和朱砂叶螨相互作用系统的研究Ⅰ.叶螨种群动态与茄子叶片单宁酸含量变化的关系. 应用生态学报, 4(2): 174-177. |
吴坤君, 李明辉. 1992. 棉铃虫营养生态学研究:食物中糖含量的影响. 昆虫学报, 35(1): 47-52. DOI:10.3321/j.issn:0454-6296.1992.01.001 |
吴坤君, 李明辉. 1993. 棉铃虫营养生态学研究:取食不同蛋白质含量饲料时的种群生命表. 昆虫学报, 36(1): 21-28. |
曾益良, 龚佩瑜, 姜立荣, 等. 1982. 施氮量对棉株和棉铃虫的影响. 昆虫学报, 25(1): 16-23. |
邹运鼎, 杨义和, 胡丽娟, 等. 1992. 两种品系油菜植株成分与蚜虫种群消长及成蚜翅型的关系. 昆虫学报, 35(2): 178-186. DOI:10.3321/j.issn:0454-6296.1992.02.012 |
Dicke M, Sabelis M W. 1988. How plants obtain predatory mites as bodyguards. Neth.J.Zool., 38: 148-165. |
Koyama K. 1985. Nutritional physiology of the brown rice planthopper Nilaparvata lugens Stal.Ⅰ.Effect of sugar on nymphal development. Appl.Ent.Zool., 20: 292-298. DOI:10.1303/aez.20.292 |
Lightfoot D C, Whitford W G. 1987. Variation in insect densities on desert creosotebush:Is nitrogen a factor?. Ecology, 68: 547-557. DOI:10.2307/1938460 |
Mattson W J. 1980. Herbivory in relation to plant nitrogen content. Ann.Rev.Ecol.Syst., 11: 119-161. DOI:10.1146/annurev.es.11.110180.001003 |
Takeshi shimoda, Junji takabayashi, Wataru ashihara, et al. 1997. Response of predatory insect Scolothrips takahashii toward herbivore-induced plant volatiles under laboratory and field conditions. J.Chem.Ecol., 23(8): 2033-2048. DOI:10.1023/B:JOEC.0000006487.49221.df |