 2002, Vol. 38
2002, Vol. 38文章信息
- 杨斌, 叶建仁, 刘吉开, 董泽军, 包宏.
- Yang Bin, Ye Jianren, Liu Jikai, Dong Zejun, Bao Hong.
- 松针褐斑病菌毒素LA-Ⅰ和LA-Ⅱ致毒活性研究
- STUDIES ON THE PHYTOTOXIC ACTIVITIES OF LA-Ⅰ AND LA-Ⅱ PRODUCING BY BROWN SPOT NEEDLE BLIGHT FUNGUS (LECANOSTICTA ACICOLA)
- 林业科学, 2002, 38(4): 84-88.
- Scientia Silvae Sinicae, 2002, 38(4): 84-88.
-
文章历史
- 收稿日期:2001-05-08
-
作者相关文章



2. 中国科学院昆明植物研究所 昆明 650204
2. Kunming Institute of Botany, The Chinese Academic of Science Kunming 650204
一种病原菌产生多种致毒性活性物质是常有的现象。这些活性物质可能因化学组成或化学结构的不同, 而在理化性质以及致病过程中的地位和作用存在明显差异(海蒂弗斯, 1991;章元寿, 1996; Pringle, 1967)。菊池链格孢菌(Alternaria kikuchiana)能产生3种毒素物质, AK Ⅰ、AK Ⅱ和TA。AK Ⅰ、AK Ⅱ是寄主专化性毒素(HST), TA为非寄主专化性毒素(NHST)。TA活性弱, 但在水稻上, TA可增强瘟病菌(Pyricularia oryzae)侵染力, 使病斑数目增加一倍, 病斑面积增大30 %。由此可见, TA有增强稻瘟病菌其他毒素活性的作用(Daly, 1983)。盘长孢刺盘菌(Colletotrichum gloeosporioides)能产生曲霉肽A和曲霉肽B(aspergillomarasmin A、B)两种毒素。曲霉肽A可引起蕃茄萎蔫并不具热稳定性; 曲霉肽B引起蕃茄叶斑, 具热稳定性。说明二者在作用机理和作用方式上明显不同(Ballio, 1972; Sharma, 1969)。正是由于多种毒素物质复杂的协同作用才造成对寄主植物的伤害。海蒂弗斯曾认为, 在植物病害过程中1种毒素很少单独起作用, 它多少总是与病原物的其他代谢产物或酶联合产生最终的致病作用(海蒂弗斯, 1991)。
松针褐斑病菌(Lecanosticta acicola)可产生毒素(海蒂弗斯, 1991)。祁高富(1999)、叶建仁(1999)曾对松针褐斑病菌毒素基本性质作过较深入研究, 认为松针褐斑病菌毒素无寄主专化性, 但不同寄主之间仍存在敏感性差异。通过半定量试验研究认为松针褐斑病菌毒素对湿地松愈伤组织的最低有效浓度为100 μ g·mL-1 (叶建仁, 1998;祁高富, 1999)。由于当时还没有分离出致病毒素, 这种试验和定量只是以松针褐斑病菌毒素粗提液全部组分为基础的。目前已经从松针褐斑病菌发酵液中分离到2个致毒活性物质LA-Ⅰ和LA-Ⅱ。根据过去其他毒素的研究结果, LA-Ⅰ和LA-Ⅱ表现出的性质和活性不一定与松针褐斑病菌毒素整体表现出来的性质与活性完全一致。在对LA-Ⅰ和LA-Ⅱ的分离纯化过程中, 为了避免丢失某些活性弱的物质, 在生测时将各分离组分均配成浓度为2 mg·mL-1的生测液进行生测(杨斌, 2001), 祁高富(1999)研究认为松针褐斑病菌毒素粗提液的致萎活性浓度应为2 mg·mL-1以下, 而目前对LA-Ⅰ和LA-Ⅱ的最低有效活性浓度还不清楚, 也不知LA-Ⅰ和LA-Ⅱ之间是否存在协同作用?二者在致病过程中地位如何?因此, 必需对LA-Ⅰ和LA-Ⅱ这2种致病毒素的基本性质进行研究, 比较二者活性大小, 研究二者的互作关系等, 为今后进一步深入研究致病毒素的致病机理奠定基础。
1 材料与方法 1.1 毒素样品根据杨斌等(2001)的方法培养、提取分离纯化获得松针褐斑病菌毒素LA-Ⅰ和LA-Ⅱ, 并经高效液相色谱检验确为纯物质。
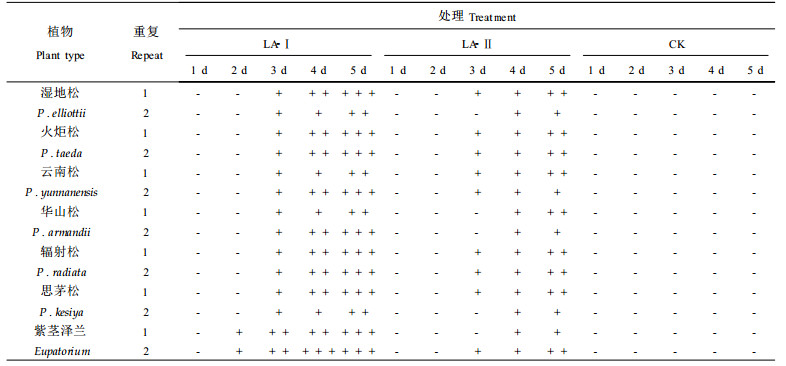
1.2 生测方法采用针刺法(叶建仁, 1998;祁高富, 1999), 以蒸馏水作为对照。生测植物材料为湿地松(Pinus elliottii) 4月生幼苗(种子由南京林业大学种子中心提供), 火炬松(P. taeda)、云南松(P. yunnanensis)、华山松(P. armandii)、思茅松(P. kesiya)和辐射松(P. radiata) 1月生幼苗(种子由云南省种苗站吴翕赠送), 紫茎泽兰(Eupatorium sp.)采自西南林学院校园。
1.3 LA-Ⅰ旋光性用旋光仪测定LA-Ⅰ旋光性, 仪器条件:光源波长λ =589 nm, 待测样品浓度为0.00947 g·mL-1, 盛液管长度20 mm, 温度13.7 ℃, 测定时先调节棱镜晶柱, 使其相互平行, 作为零度, 将被测样品溶液(以CH3OH作溶剂)放入盛液管, 测量旋光度。
1.4 LA-Ⅰ、LA-Ⅱ热稳定性LA-Ⅰ、LA-Ⅱ配成浓度为1 mg·mL-1溶液, 置于沸水中加热15 min, 冷却后生测, 以不加热的LA-Ⅰ和LA-Ⅱ为对照, 生测材料为湿地松幼苗、紫茎泽兰叶片。
1.5 LA-Ⅰ、LA-Ⅱ专化性将LA-Ⅰ、LA-Ⅱ配成浓度为2 mg·mL-1水溶液, 用不同松树幼苗、紫茎泽兰叶片为生测材料。
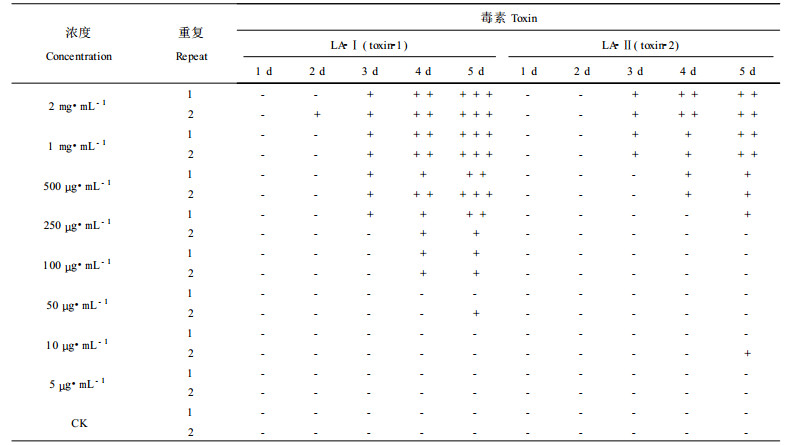
1.6 LA-Ⅰ、LA-Ⅱ最低有效浓度将LA-Ⅰ、LA-Ⅱ配在浓度为2 mg·mL-1、1 mg·mL-1、500 μ g·mL-1、250 μ g·mL-1、100 μ g·mL-1、50 μ g· mL-1、10 μ g·mL-1、5 μ g·mL-1水溶液, 进行生测, 生测材料为湿地松幼苗, 以蒸馏水为对照(最低有效浓度指出现明显伤害症状的最低毒素浓度)。
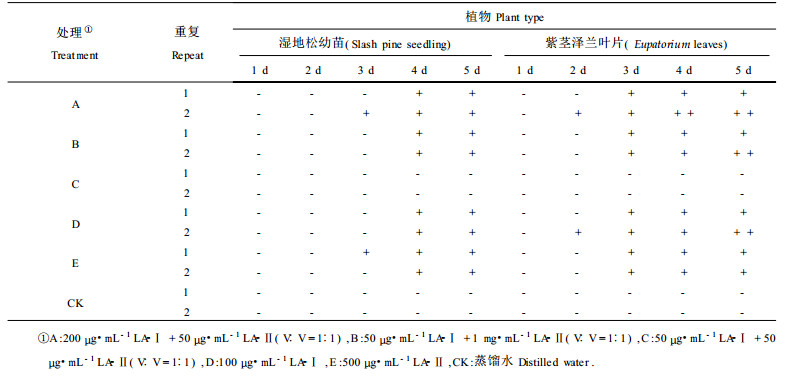
1.7 LA-Ⅰ、LA-Ⅱ相互关系用200 μ g·mL-1LA-Ⅰ +50 μ g·mL-1LA-Ⅱ(体积比为1: 1), 50 μ g·mL-1LA-Ⅰ +1 mg·mL-1LA-Ⅱ(体积比为1: 1), 100 μ g·mL-1LA-Ⅰ, 500 μ g·mL-1LA-Ⅱ溶液进行生测, 观察LA-Ⅰ、LA-Ⅱ之间是否有相互促进或相互抑制的现象, 以水为对照, 生测材料为湿地松幼苗。
2 结果与分析松针褐斑病菌毒素LA-Ⅰ的分子结构为HOOCCH(CH3) OCH(CH3) COOH, 具2个手性碳原子1)。旋光测定结果显示LA-Ⅰ旋光度α=+0.0010°, 按公式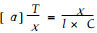
1) 杨斌.松针褐斑病菌致病毒素的研究.南京林业大学.博士学位论文, 2002, 6.
|
|
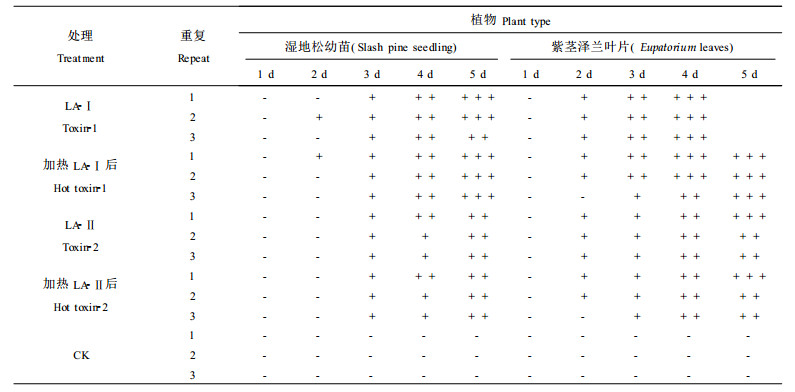
LA-Ⅰ和LA-Ⅱ不仅能为害湿地松、火炬松和辐射松幼苗, 而且对云南松、思茅松和华山松幼苗也有伤害作用。在浓度为2 mg·mL-1条件下生测, LA-Ⅰ和LA-Ⅱ对以上几种松树幼苗的伤害程度无明显差异。伤害症状多为松苗出现萎蔫, 有时可见明显水渍状段斑和黄色段斑。而用毒素原液或毒素粗提液生测时, 几乎每个生测材料上均出现段斑。LA-Ⅰ和LA-Ⅱ对杂草紫茎泽兰也有伤害, 其伤害症状与毒素粗提液处理时一致, 即在叶片上出现红褐色小点或出现近圆形水渍状斑。LA-Ⅰ和LA-Ⅱ为非寄主专化性毒素, 供试的几种松树的松苗对这两种毒素敏感性差异不明显(见表 2)。
|
|
研究表明LA-Ⅰ最低有效浓度为100 μ g·mL-1, LA-Ⅱ最低有效浓度为500 μ g·mL-1。LA-Ⅰ的活性比LA-Ⅱ高5倍左右。100 μ g·mL-1的LA-Ⅰ在5 d内可使生测材料有明显枯斑, 但不能使整个针叶全部枯死或萎蔫; 而当浓度为500 μ g·mL-1时, 在5 d内就可使大部分生测材料枯死或萎蔫。LA-Ⅱ活性弱一些, 浓度要在1 mg·mL-1时才可使生测材料在5 d内全部枯死(见表 3)。
|
|
为了灵敏地观察LA-Ⅰ和LA-Ⅱ之间的相互作用, 试验选择了最低有效浓度作为生测浓度。研究发现, 在含量低于有效浓度的LA-Ⅰ和LA-Ⅱ中分别加入少量LA-Ⅰ和LA-Ⅱ, 生测活性均无明显变化; 低于最低有效浓度的LA-Ⅰ和LA-Ⅱ混合对生测材料没有伤害作用。由此可见, LA-Ⅰ和LA-Ⅱ之间不存在明显相互增强或削弱活性的情况(表 4)。
|
|
植物病原菌毒素根据其对植物的特异性作用不同, 可分为寄主专化性毒素(HST)和非寄主专化性毒素(NHST)。寄主专化性毒素致病的植物范围与产毒病菌的寄主范围一致。非寄主专化性毒素除能对寄主植物产生毒害作用外, 对非寄主植物也有毒害作用。本研究用提纯后的毒素LA-Ⅰ和LA-Ⅱ进行生测, 表明这两种毒素不仅对湿地松、火炬松、辐射松、云南松和华山松等有伤害作用, 而且对草本植物紫茎泽兰也有伤害作用。说明LA-Ⅰ和LA-Ⅱ为非寄主专化性毒素。叶建仁等曾用毒素粗提液对多种植物进行生测, 也显示粗毒素有非寄主专化性的特点(海蒡弗斯, 1991)。但不同植物对毒素粗提液的敏感性还存在种间差异。从本试验生测结果看, 以LA-Ⅰ、LA-Ⅱ进行生测, 这种种间差异表现不十分明显。随着对各种植物病原菌毒素的深入研究, 人们逐渐发现一种病原菌往往产生多种致毒物质来适应不同寄主植物和同一寄主植物的不同抗病机制, 而且多种毒素组分协同作用才对植物造成伤害。LA-Ⅰ、LA-Ⅱ专化性试验结果与过去用毒素粗提液进行专化性研究的结果基本一致但并非完全相同, 其原因可能是毒素粗提液中除了含有LA-Ⅰ和LA-Ⅱ活性组分外还有其他的化合物, 影响了毒素的活性和种间选择性。
过去用半定量方法研究毒素作用的最低有效浓度表明:松针褐斑病菌毒素对湿地松愈伤组织的最低有效浓度在100 μ g·mL-1以下。本试验用湿地松幼苗作生测材料进行最低浓度研究表明, LA-Ⅰ活性比LA-Ⅱ活性高4 ~ 5倍, LA-Ⅰ作用的最低有效浓度为100 μ g·mL-1, LA-Ⅱ为500 μ g·mL-1, 若以愈伤组织为生测材料, 这数字应当要小得多。同其他植物病原菌毒素的作用浓度相比, 松针褐斑病菌毒素LA-Ⅰ和LA-Ⅱ的活性并不高, 过去发现的毒素物质一般在浓度为10 μ g·mL-1就表现出很强的致毒活性。LA-Ⅰ和LA-Ⅱ互作关系研究表明, LA-Ⅰ和LA-Ⅱ之间没有相互促进, 也没有明显相互抑制的现象。LA-Ⅱ是一种含S化合物(杨斌, 2001), 根据Youla对含硫化合物的生物活性和功能的研究, 认为含S活性物质通常有加强或模仿其他生物活性物质的功能。松针褐斑病菌毒素中是否存在其他含量极低的活性物质与LA-Ⅱ协同作用值得进一步研究。
海蒂弗斯R, 威廉斯P H主编, 朱有红, 宋佐衡, 傅淑云等译.植物病理生理学.北京: 农业出版社, 1991
|
祁高富, 叶建仁. 1999. 松针褐斑病菌毒素的确定及其基本性质的研究. 南京林业大学学报, 23(4): 17-21. DOI:10.3969/j.issn.1000-2006.1999.04.004 |
杨斌, 刘吉开, 叶建仁等.松针褐斑病菌毒素LA-Ⅱ的分离纯化及化学结构研究.南京林业大学学报, 2001, 25(3): 章元寿主编.植物病理生理学.南京: 江苏科学技术出版社, 1996 http://www.cnki.com.cn/Article/CJFDTotal-LYKE200502013.htm
|
叶建仁, 祁高富. 1999. 松针褐斑病菌毒素的专化性研究. 南京林业大学学报, 23(6): 1-4. DOI:10.3969/j.issn.1000-2006.1999.06.001 |
叶建仁, 解春霞. 1998. 松针褐斑病菌致病机制研究. 林业科学研究, 11(3): 243-248. DOI:10.3321/j.issn:1001-1498.1998.03.003 |
Ballio A. Phytotoxins: a exercise in the chemistry of biologically active natural products.In : Phytotoxins in Plant Disease, New York-London : Academic press, 1972
|
Daly J M, Deverall B J. 1983. Toxins and plant pathogenesis. New York-London: Academic press.
|
Hawes M C. 1983. Technique for using isolated corn root loop cells in a simple, quantative bioassay for the pathotoxin produced by Helminthosporium maydis Race T. Phytopathology, 73: 1184-1187. DOI:10.1094/Phyto-73-1184 |
Pringle R B, Scheffer R P. 1967. Multiple host-specific toxins from Periconia circinata. Phytopathology, 57: 530-532. |
Sharma M C, Sharma B C. 1969. Toxic metabolite production by colletotrichium gloeosporioides causing citrus die-back in India. Phytopathology, 22: 67-74. |