 2002, Vol. 38
2002, Vol. 38文章信息
- 李建民, 周志春, 吴开云, 金国庆.
- Li Jianmin, Zhou Zhichun, Wu Kaiyun, Jin Guoqing.
- RAPD标记研究马褂木地理种群的遗传分化
- GENETIC DIFFERENTIATION OF GEOGRAPHIC POPULATIONS IN LIRIODENDRON CHINENSE USING RAPD MARKERS
- 林业科学, 2002, 38(4): 61-66.
- Scientia Silvae Sinicae, 2002, 38(4): 61-66.
-
文章历史
- 收稿日期:2001-11-19
-
作者相关文章


2. 中国林业科学研究院亚热带林业研究所 富阳 311400
2. The Research Institute of Subtropical Forestry, CAF Fuyang 311400
种群是进化的基本单位, 种群遗传结构则是一个物种最基本的特征之一。因自然或人为原因发生突变、基因流、选择和遗传漂变, 使种群间产生遗传分化。研究种群间的遗传分化及影响因子是群体遗传学的重要课题, 是探讨植物适应性、物种形成过程及其进化机制的基础, 也是保护生物学和遗传资源学的核心(邹喻萍等, 2001)。
马褂木(Liriodendron chinense)是木兰科孑遗植物, 为我国二类保护树种。由于第四纪冰川的作用, 马褂木在地理上呈“一带五岛”分布形式, 间断星散分布于我国亚热带的广大区域(郝日明等, 1995)。生境片断化, 加之其特有的致濒机制及长期人为的选择压力, 对马褂木地理种群的遗传发化产生巨大作用。种源试验结果表明, 马褂木主要经济性状在地理种群间存在显著的遗传差异, 并呈随机变异模式, 即使两相邻种群也产生巨大的分化(李建民, 2001; 李斌等, 2001)。虽然罗光佐等(2000)曾利用RAPD标记对马褂木和北美鹅掌揪的种间遗传多样性进行了分析和比较, 但仅涉及6个马褂木地理种群。本文基于种源试验结果, 利用RAPD标记研究马褂木全分布区地理种群间的遗传分化及变异模式, 并在DNA水平上区划马褂木地理种群, 从而为有效保护和开发利用马褂木优良种质资源提供理论指导。
1 材料与方法 1.1 试验材料试验叶样取自设置在福建省邵武市的8 a生马褂木全分布区种源试验林, 其中马褂木参试种源有安徽黄山、大别山, 浙江松阳, 江西庐山、黄冈山, 湖北恩施, 福建武夷山, 湖南绥宁、桑植、浏阳, 贵州松桃、黎平, 四川叙永、酉阳, 云南蒿明15个。2001-05, 在种源试验林第1重复内, 对每种源随机采集8个不同单株的嫩叶用于DNA提取。
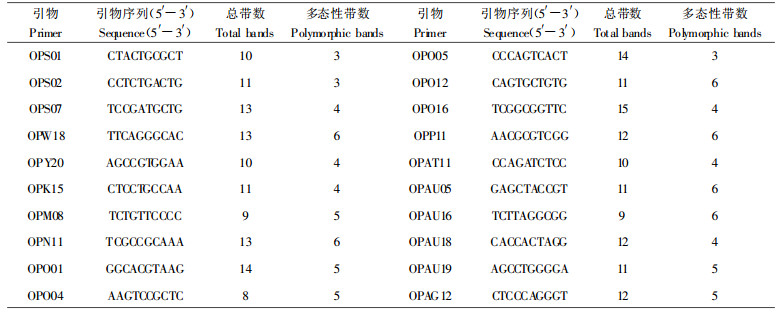
1.2 样品处理新鲜嫩叶5 g采用CTAB法提取DNA(Doyle et al., 1987)。参照李周岐1)对杂交马褂木的研究结果筛选20个能产生多态性、清晰明亮谱带的引物。各引物名称、序列及检测的总带数和多态性带数详见表 1。电泳图的标记量为λDNA/EcoR Ⅰ +Hind Ⅲ Marker。
|
|
1) 李周岐.鹅掌楸属种间杂种优势的研究。南京林业大学博士学位论文, 2000.
RAPD扩增反应在Perkin-Elmer 9600基因扩增仪上进行。RAPD扩增反应条件:94 ℃预变性3 min; 然后进入45个循环, 每个循环94 ℃变性30 s, 36 ℃退火30 s, 72 ℃延伸1.5 min; 循环完成后于72 ℃延伸7 min, 最后于4 ℃下保存。反应采用25 μL体系:10倍反应缓冲液2.5 μL(100 mmol Tris-HCl pH9.0, 500 mmol KCl, 25 mmol MgCl2, 0.01%明胶, 5.0 g·L-1 BSA), Taq聚合酶1单位, dATP、dTTP、dCTP和dGTP各100 μmol, 引物5 pmol, DNA模板50 ng左右。通过研究比较, 扩增产物在1%含溴化乙锭的琼脂糖凝胶中电泳分离, 最后在紫外灯下用Polaroid摄像系统拍照记录结果(图 1)。

|
图 1 引物OPS01的RAPD扩增产物在部分马褂木种源植株样品中的分离 Fig. 1 Segregation of RAPD bands amplified with primer OPS01 in some provenance samples of L.chinense M:分子量标记Molecular weight marker; 3-7~5-2:马褂木种源样品L.chinense provenances under analyzing. |
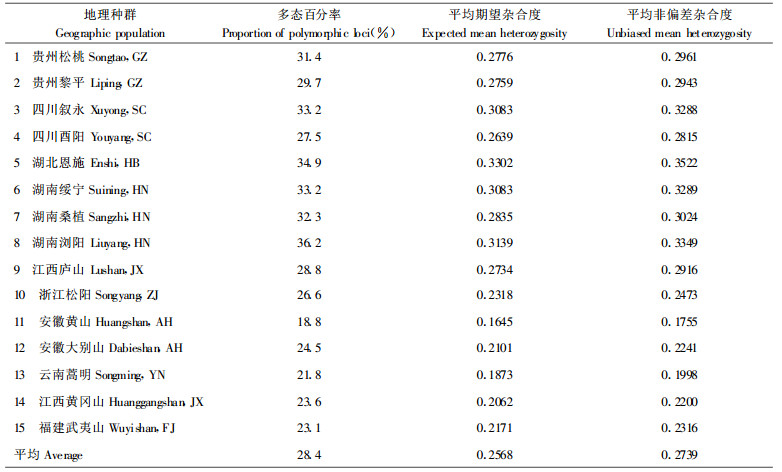
RAPD谱带按1/0标记, 利用TFPGA(Tools for Population Genetic Analyses)软件(Version 1.3)进行数据的遗传分析。首先计算衡量种群遗传变异的多态百分率(Proportion of polymorphic loci)、平均期望杂合度(Expected mean heterozygosity)和平均非偏差杂合度(Unbiased mean heterozygosity), 以度量马褂木各地理种群的遗传变异。采用F-统计量分析种群的基因多样性, 估算基因分化系数和固定系数, 以研究确定马褂木种群间的遗传分化。基于地理种群间Ner's(1978)无偏遗传距离(Unbiased genetic distance)的估算, 用非加权成组配对法(UPGMA)对15个地理种群进行聚类, 构建聚类树状图, 据此在DNA水平上对马褂木进行种源区划。
2 结果与分析参照李周歧1)筛选的20个引物对15个马褂木地理种群进行随机扩增, 发现在总共229个检测位点中有94个位点呈多态性。
1) 李周岐.鹅掌楸属种间杂种优势的研究.南京林业大学博士学位论文, 2000.
多态百分率、平均期望杂合度或平均非偏差杂合度是检验种群内遗传多样性的常用指标, 尤其是后两者不依赖于人为定义, 从基因频率的角度度量种群的遗传多样性。从表 2可见, 马褂木不同种群间的多态百分率差异巨大, 平均多态百分率为28.4%, 湖南浏阳种群高达36.2%, 而安徽黄山种群仅为18.8%, 相差1倍左右。从平均非偏差杂合度估算值来看, 马褂木种群具有较为丰富的遗传多样性, 其种群平均值为0.2739, 高于一般针阔叶树种0.206的估算值(葛颂, 1988)。然而种群间杂合度相差巨大, 多数种群的遗传多样性丰富, 但诸如安徽黄山、云南蒿明等种群内的遗传变异水平较低。与地理径纬度的相关分析表明, 种群的遗传多样性与地理纬度无关, 似与地理径度有关。种群的3个遗传多样性指数与地理径度的相关系数为-0.40, 东部种群的遗传多样性较低, 而西部种群的遗传多样性较高。
马褂木是古老的残遗植物, 虽然在我国亚热带地区都有分布, 但呈星散间断分布, 除西部种群外, 多数种群的个体数较小, 在1 ~ 20株之间(郝日明等, 1995)。这种生境片断化、小种群效应和距离隔离效应的结果, 应使得马褂木种群内的遗传变异程度降低, 而种群间的遗传分化程度加大。基因分化系数(GST)和固定指数(FST)估算结果显示, 马褂木有30%以上的遗传变异存在于种群间, 而小于70%以上的遗传变异来自于地理种群内, 这一估算值近似于我国特有濒危植物银杉(Cathaya argyrophylla)的研究结果(汪小全等, 1996; Ge et al., 1998), 与朱晓琴等(1995)和罗光佐等(2000)对马褂木的研究结果相反。罗光佐等(2000)利用RAPD标记研究6个马褂木种群和5个北美鹅掌楸种群的遗传多样性(表 3), 结果认为马褂木的遗传变异主要来自地理种群内而非地理种群间, 这难以从本质上解释马褂木因生境长期片段化和以小种群繁育形式而出现的种群遗传分化, 与马褂木种源试验结果不符。
|
|
|
|

表 4给出了马褂木15个地理种群间Nei's(1978)非偏差遗传距离, 并据此使用UPGMA聚类法得出树状图。图 2聚类树状图显示, 利用RAPD标记可以很好地按地理区域将马褂木划分成两个种源区, 第1种源区主要包括马褂木分布区的中、西部种群, 这是马褂木的主要分布中心, 其种群一般较大, 种群间的遗传分化较小。如四川酉阳、湖北恩施和湖南绥宁3个种群相隔不远, 其遗传距离也不大, 在0.0310 ~ 0.0994之间。同样相距不远的湖南桑植、浏阳和江西庐山种群其遗传距离也仅在0.0476 ~ 0.0972的范围内。第1种源还可区划成2个种源亚区, 即马褂木西南亚区和华中亚区(不完全是行政上的华中地区), 前者包括四川叙永、酉阳, 湖北恩施和湖南缓宁, 贵州松桃、黎平等种群, 后者包括湖南桑植、浏阳和江西庐山种群。根据聚类树状图, 马褂木第2种源区主要包括福建、浙江和安徽等分布区。在这一种源区中马褂木分布稀少, 种群很小, 其结果种群间的遗传分化较之第1种源区更大, 如位于武夷山山脉东西两侧, 相距仅数十公里的福建武夷山和江西黄冈山两个种群, 遗传距离达0.1369。有一点难以理解的是, 在聚类分析中地域上不相连的云南蒿明也划在第2种源区中。
|
|


|
图 2 15个地理种源UPGMA聚类法得出的树状图 Fig. 2 Dendrogram showing Nei's unbiased genetic distance among 15 populations using UPGMA cluster analysis |
根据化石资料, 在新生代时鹅掌楸属有10多种, 分布甚广, 中欧、北美及格陵兰上白垩纪地层中都曾发现可靠的化石。新生代初, 曾生存于冰岛; 中世纪时, 曾生存于瑞士与日本; 上新世时在欧洲的法国、荷兰及意大利更为茂盛, 中亚一带的阿尔泰山脉, 在上新世地层中也曾发现化石; 第四纪冰期以后, 鹅掌楸在欧洲及其它各处均已绝灭, 仅存马褂木和北美鹅掌楸两种(王明庥, 2001)。马褂木现残遗和间断分布于我国南方各省区, 其中以贵州、湖南和湖北为分布中心, 东南诸省虽有分布, 但种群很小。利用RAPD标记对马褂木全分布区15个种群进行了遗传多样性分析, 结果显示, 马褂木虽因小种群效应和距离隔离效应经历了严重的遗传漂变, 种群内仍具有丰富的遗传变异, 但东部种群的遗传多样性偏低, 这符合小种群的遗传结果。基因分化系数和固定指数揭示, 由于小种群效应, 以及缺乏有效的基因流和马褂木特有的致濒机制, 马褂木种群间的遗传分化巨大, 30%以上的遗传变异存在于种群间, 与我国濒危树种银杉(汪小全等, 1996; Ge et al., 1998)、红桧和台湾扁柏(黄士颖等, 1998)相似, 而与广布性树种不同。朱晓琴等(1995)和罗光佐等(2000)曾分别利用等位酶和RAPD标记研究马褂木的遗传多样性, 认为仅有低于10%的遗传变异来自马褂木种群间, 这不能解释马褂木生境片断化和小种群的遗传结果。
作者曾根据单个地点的种源试验测定材料将这15个种群进行聚类, 虽然也同样将马褂木分成东部和西部两个种源区, 但有不少种源难以正确的归类(李建民, 2001)。罗光佐等(2000)基于种群RAPD指纹图谱的相似性, 虽能将马褂木与北美鹅掌楸很好地区分开来, 但难以将浙江、贵州、江西、湖北、湖南和四川6个马褂木种群按地域归类。本文利用RAPD标记在DNA水平进行种源区划, 可以很好地将马褂木划分为西部和东部两个种源区, 而西部种源又可划分为西南亚区和华中亚区。西部种源区是马褂木的分布中心, 现有种群较大, 在地域上相距不远的种群其遗传距离较小, 但在东部种源区中由于现有的种群都较小, 即使两个相邻种群, 其遗传距离也非常大。
葛颂. 1988. 同功酶与林木群体遗传变异研究. 南京林业大学学报, 12(1): 68-77. |
郝日明, 贺善安, 汤诗杰, 等. 1995. 鹅掌楸在中国的自然分布及其特点. 植物资源与环境, 4(1): 1-6. |
黄士颖, 林惠文, 郭怡秀等.以逢机扩增多型性DNA分子标记探讨红桧与台湾扁柏遗传变异.两岸林木种源交流研讨会论文集(台湾省林业试验所林业从刊第88号), 1998, 137~144
|
李斌, 顾万春, 夏良放, 等. 2001. 鹅掌楸种源遗传变异和选择评价. 林业科学研究, 14(3): 237-244. DOI:10.3321/j.issn:1001-1498.2001.03.001 |
李建民. 2001. 马褂木地理遗传变异和优良种源选择. 林业科学, 37(4): 41-49. DOI:10.3321/j.issn:1001-7488.2001.04.007 |
罗光佐, 施季森, 尹佟明, 等. 2000. 利用RAPD标记分析北美鹅掌楸与鹅掌楸种间遗传多样性. 植物资源与环境学报, 9(2): 9-13. DOI:10.3969/j.issn.1674-7895.2000.02.003 |
汪小全, 邹喻萍, 张大明. 1996. 松科系统发育的分子生物学证据. 植物分类学报, 35(2): 97-106. |
王明庥. 2001. 林木遗传育种学. 北京: 中国林业出版社, 73.
|
邹喻萍, 葛颂, 王晓东. 2001. 系统与进化植物学中的分子标记. 北京: 科学出版社.
|
朱晓琴, 马建霞, 姚青菊, 等. 1995. 鹅掌楸遗传多样性的等位酶论证. 植物资源与环境, 4(3): 9-14. |
Doyle J J, Doyle J L. 1987. A rapid DNA isolation procedure for small quantities of fresh leaf material. Phytochem Bull, 19: 11-15. |
Ge S, Hong D Y, Wang H Q, et al. 1998. Population genetic structure and conservation of an endangered conifer, Cathaya argyrophylla(Pinaceae). International Journal of Plant Science, 159: 351-357. DOI:10.1086/297556 |
Nei M. 1978. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics, 89: 583-590. |