 2002, Vol. 38
2002, Vol. 38文章信息
- 冯建灿, 张玉洁.
- Feng Jiancan, Zhang Yujie.
- 喜树光合速率日变化及其影响因子的研究
- STUDIES ON THE DIURNAL CHANGES OF NET PHOTOSYNTHESIS RATE AND THE EFFECT OF ENVIRONMENTAL FACTORS OF CAMPTOTHECA ACUMINATA
- 林业科学, 2002, 38(4): 34-39.
- Scientia Silvae Sinicae, 2002, 38(4): 34-39.
-
文章历史
- 收稿日期:2001-10-18
-
作者相关文章


2. 河南省林业技术推广站 郑州 450008
2. Forestry Technique Extending Station of Henan Province Zhengzhou 450008
喜树(Camptotheca acuminata)属珙桐科(Nyssaceae)旱莲属植物。主要分布于我国长江流域及西南各省和印度部分地区, 我国台湾、浙江、广西、河南等地也有栽培。1966年美国的Wall从喜树皮中分离出喜树碱(Camptothecin, CPT), 经肿瘤试验证明这种色氨酸-萜烯类生物碱具有抗癌活性(Wall, 1966), 从而引起了人们的广泛关注。1985年, Hsiang发现CPT能阻断拓朴异构酶Ⅰ (topoisomerase Ⅰ)的合成(Hsiang, 1985), 而不是拓朴异构酶Ⅱ(topo Ⅱ), 这正是CPT独特的抗癌机理(Hsiang, 1988)。这一发现掀起了喜树与CPT研究的新高潮。CPT还被发现能阻断HIV和其它逆转录酶病毒如马贫血病病毒和鼠白血病病毒等在培养细胞内的复制(Priel, 1991)。由于CPT药用价值, 使喜树成为红豆杉之后第二个重要的木本抗癌药用植物, 已成为世界性热门研究课题。
国内对喜树和喜树碱也进行了多方面的研究, 并取得了可喜的研究成果(冯建灿, 2000)。但对喜树生理特性如光合特性、蒸腾特性及环境因子对这些生理反应的影响等方面的研究尚未见报道。本文对喜树的光合特性及其与主要环境因子间的关系进行了探讨, 以期为喜树药用林栽培提供理论指导。
1 材料与方法 1.1 材料试验材料为2 a生喜树幼树。种子采自河南鸡公山, 1998年春播种, 由于苗木顶部受冬季低温和春季干旱等影响而抽条, 于1999-03初将干枯部分剪除。
1.2 测定方法试验从1999-05开始在河南农业大学林学园艺学院试验站内进行, 10月份结束。从5月初起, 每10 d左右选择一晴天进行1次日变化测定。测定时间为每天7:00到18:00, 每间隔1 h测定1次。
在苗冠中部选择2~3片成熟叶进行测定, 每叶片重复5次, 每时段测定4株。
测定仪器为美国CID公司生产的CI-301型便携式CO2气体分析仪, 开放气路测定。
仪器自动记录净光合速率(Pn)、大气温度(Tair)、叶片温度(TLeaf)、叶室进口相对湿度(RHin)、叶室出口相对湿度(RHout)、大气CO2浓度(CO2 in)、叶室出口CO2浓度(CO2 out)、细胞间隙CO2浓度(CO2 int)、PFD(photon flux density), 气孔导度(C)和蒸腾强度(E)等(文中描述亦用此缩写)。
所测得数据输入计算机进行统计模拟与计算, 计算方法见张贤珍(1990)、丁希泉(1989)、郎奎建(1989)和SPSS(SPSS Inc. 1998)数据统计软件包。
2 结果与分析 2.1 晴天叶片光合速率与光合量子效率日变化喜树Pn日变化如图 1所示。晴天时Pn日变化为不对称的双峰曲线, 最大值出现在上午10:00前后, 为7.9075 μmolCO2·m-2s-1。此后Pn持续下降, 到14:00左右出现低谷值2.4470 μmolCO2·m-2 s-1。而后Pn缓慢回升, 直到16:00至17:00又出现第二个峰值, 为3.7340 μmolCO2·m-2 s-1, 升高幅度为1.287 μmolCO2·m-2s-1, 保持短时间的高峰后迅速下降。比较两个峰值发现, 下午的峰值仅为上午峰值的1/2。一天中Pn的极差达8.25 μmolCO2·m-2s-1。

|
图 1 喜树叶片光合速率日变化 Fig. 1 Diurnal changes of Pn in leaves of C. acuminata
 Pn晴Clear; Pn晴Clear;  Pn阴Cloudy; 下同.Same in below. Pn阴Cloudy; 下同.Same in below.
|
阴天时, 喜树Pn日变化为一单峰曲线, 上午逐渐上升, 下午不断下降, 峰值出现在12:00前后, 为5.2 μmolCO2·m-2s-1, 仅为晴天上午峰值的65.76 %。日平均光合速率为3.75 μmolCO2·m-2s-1, 仅为晴天时4.33 μmolCO2·m-2 s-1的85 %。
2.2 光合速率与主要环境因子的关系图 2至图 4所示, 在晴天10:00, PFD即高达1121.6 μmol·m-2 s-1, 低于光饱和点(数据另文发表), Pn达到第一个峰值, 之后随着PFD增强, 气温升高, 空气相对温度急剧下降, Pn也迅速下降, 14:00降到低谷。然后PFD、气温开始下降, 相对湿度逐步升高, Pn也逐步出现午后高峰(16:00)。16:00后, PFD和气温继续降低, 相对湿度快速升高, Pn也迅速降低。

|
图 2 喜树叶片光合速率与光通量密度间关系 Fig. 2 Relationship between diurnal changes of Pn and PFD
 PFD晴Clear; PFD晴Clear;  PFD阴Cloudy PFD阴Cloudy |

|
图 3 喜树叶片光合速率与气温间关系 Fig. 3 Relationship between diurnal changes of Pn and air temperature
T晴Clear; T阴Cloudy |

|
图 4 喜树叶片光合速率与相对湿度关系 Fig. 4 Relationship between diurnal changes of Pn and relative humidity
RH晴Clear; RH阴Cloudy |
在阴天时, 由于PFD始终低于喜树的光饱和点, 因此Pn变化与PFD和气温的变化一致, 与相对湿度等因子的变化方向相反。
经过夜间的富集, 大气中CO2浓度在早晨最高(图 5), 日出后, 随着植物光合作用的进行, CO2浓度逐步降低, 11:00以前大气中CO2浓度下降的速度很快, 到11:00降至一低谷值251.1 μL·L-1, 较早晨降低了87.7 μL·L-1; 此后有所回升, 14:00左右CO2浓度则有一个午间高峰值出现, 16:00出现日间最低值238.45 μL·L-1。大气中CO2浓度与Pn间的关系较为复杂, 基本表现为随着Pn的增大, 大气中CO2浓度逐渐降低, 随着Pn降低而增大。值得注意的是在午间, 当午间Pn出现低谷值时, CO2浓度的则为午间峰值260.6 μL·L-1(14:00)。这是由于午间光合速率降低, 形成的CO2聚集。

|
图 5 喜树叶片光合速率与CO2浓度间关系 Fig. 5 Relationship between Pn and CO2 concentration in air
CO2晴Clear; CO2阴Cloudy |
阴天时Pn与大气中CO2的关系十分清楚, 午前随着Pn的增大, CO2逐渐降低, 午后则相反。
2.3 Pn与气孔导度和细胞间CO2浓度间的关系晴天时, 喜树叶片气孔导度的日变化为双峰型(图 6), 高峰值出现在8:30前后, 气孔导度为429.9 mmolH2O·m-2s-1, 午后的峰值出现在15:00左右, 仅为142.4mmolH2O·m-2s-1, 而且高峰不太明显。气孔导度与Pn的变化基本是平行的, 两者最大值的相差约为2 h, 但最低值出现的时间相同, 表明Pn的变化落后于气孔开张程度的变化。

|
图 6 喜树叶片光合速率与气孔导度关系 Fig. 6 Relationship between Pn and stomatal conductance in leaves of C. acuminata
C晴Clear; C阴Cloudy |
由图 7可以看出, 从10:00~11:00气孔导度开始逐渐下降, 细胞间CO2浓度也同步下降, Pn也逐渐下降, 它们的变化趋势是一致的, 由此可以认为在这段时间内, Pn的变化主要是由气孔运动来进行控制。此后气孔导度下降则引起细胞间CO2浓度的上升, 而不是下降, 这种现象一直持续到日落。依据Farquhar和Sharkeyr的观点, 细胞间CO2供应不是导致Pn下降的直接原因, 这段时间Pn的变化主要是由非气孔因素控制, 如1, 5-二磷酸羧化酶(Rubisco)活性与含量等因素。

|
图 7 喜树叶片光合速率与细胞间CO2关系 Fig. 7 Relationship between Pn and the intercellular CO2 concentration in leaves of C. acuminata
CO2int晴Clear; CO2int阴Cloudy |
阴天时, 午前随着Pn的增大CO2 int逐渐降低, 而午后则相反, CO2 int保持在较高的水平上。这是由于在阴天条件下, 温度和光照等条件不能满足光合机构进行光合作用的要求, 进入叶内的CO2没有被固定形成的积累所致。
2.4 Pn与蒸腾强度间的关系如图 8所示, 无论是晴天还是阴天, 蒸腾强度的日变化均为一单峰曲线。晴天时, 在Pn达到峰值时, 蒸腾强度为6.4271 mmolH2O·m-2s-1, 接近高峰值, 但其高峰值6.9635 mmolH2O·m-2s-1出现在午间, 午后蒸腾强度则持续下降, 两者的变化不一致。阴天时Pn与蒸腾强度的升降完全同步。

|
图 8 喜树叶片光合速率与蒸腾强度关系 Fig. 8 Relationship between diurnal changes of Pn and transpiration rate
E晴Clear; E阴Cloudy |
从以上分析不难看出主要环境因子与C、CO2 int和E等内在因子均对Pn产生了一定程度的影响, 为进一步分析其间的数量化关系, 对测定结果进行了回归分析。
Pn与这些因子间的数量关系由多元线性方程分别表示为:
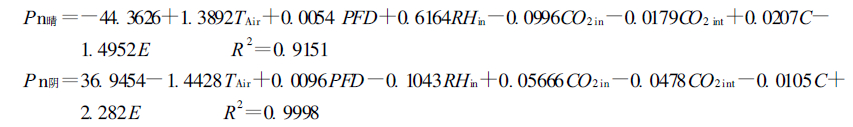
|
由决定系数R2可以看出, 这些因子是影响Pn的重要因子, 但其所起的作用不同。与Pn的相关系数由大到小的顺序为C、E、PFD、TAir、CO2 in、CO2 int、RHin。
|
|

|
|
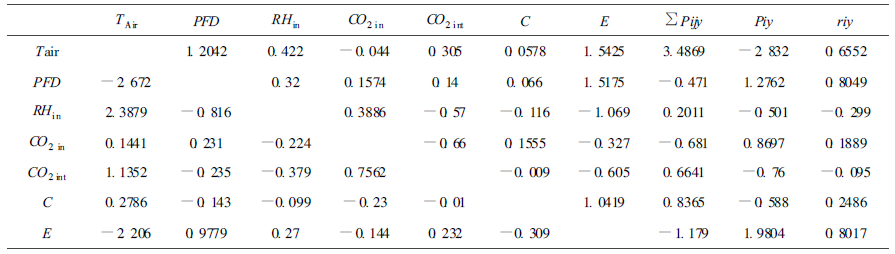
通径分析结果表明:晴天时, 主要因子对Pn的直接影响系数由大到小顺序为RHin、TAir、CO2 in、E、PFD、C和CO2 int, 气孔限制的作用在晴天Pn日变化中所起的作用较小。在相关分析中显得不为重要的相对湿度和气温上升到了重要位置, 这是由于此2个因子间存在着极强的负相关性(-0.9451), 使它们对Pn的影响在相关分析中被抵消, 从表中的间接通径系数亦可看出。由此可以认为, 晴天时对Pn影响最大的因子是RHin, RHin每升高一个标准单位, Pn可提高3.9133个标准单位。这与喜树原产长江流域, 长期适应该地区湿热的环境条件有关。引种到北方后, 在重要的生长时节, 如何提高其生长环境的水热条件, 应成为制定栽培技术措施的关键。
从表 2简单相关分析结果不难看出, 在阴天情况下只有3个因子与Pn的相关达到了显著水平, PFD和TAir与Pn间的相关关系达到了极显著水平; E与Pn间的相关关系达到了显著水平, 这一方面说明Pn和E日变化的一致性, 同时表示出光合作用与蒸腾之间存在紧密的联系。RHin和CO2 in与Pn的相关关系没有达到显著水平。由此可以认为, 在阴天情况下, 由于光照不足, 使PFD成为限制光合作用的重要因子, 这从另一角度证明喜树是典型的阳性植物。
通径分析结果与简单相关的结果基本一致, TAir、E和PFD对Pn的影响较大。结合上述分析不难得出PFD是影响阴天喜树Pn的关键因子, 虽然其直接通径系数小于TAir, 但TAir的变化受制于PFD的强弱。
值得注意的是在阴天条件下, C的直接和间接通径系数均较小。
3 结论与讨论植物光合速率日变化及其影响因素一直是科技工作者研究的焦点之一, 已有报道对茌梨(苗德全, 1991)、核桃(张志华, 1993)、银杏(陶俊, 1999)、香椿(徐坤, 1999)、苹果(程来亮, 1992)、野生毛葡萄(朱林, 1994)、猕猴桃(彭永宏, 1994; 刘旭峰, 1993)等多种经济树种进行了研究, 草本植物方面的研究就更多。郑丕尧(1992)将植物的晴天Pn日变化分为单峰型、双峰型、严重型和平坦型四种类型。在晴天时喜树Pn日变化为双峰曲线, 属严重型, 具有典型的“午休”特征。阴天时则为单峰型。
影响喜树叶片Pn日变化的主要生态因子是PFD、相对湿度, 由于光强的不易控制性, 在生产中可通过浇水、增加相对湿度等措施提高喜树的光合速率。
在阴天条件下PFD是影响喜树Pn的关键因子, 虽然其直接影响小于气温, 但气温的变化受制于PFD的强弱。这也证明了喜树的喜光性。
对于植物Pn产生“午休”内在原因的研究结果极不相同。由于气孔是CO2和水分进出叶片的重要通道, 其开张程度或气孔阻力的大小, 对植物的水分状况和CO2同化有着重要的影响。一般认为气孔开度减小即阻力增大, CO2进入叶片受阻, CO2int降低, Pn下降; 同时气孔阻力也减少, 水分的散失也蒸腾速率也下降, 得出这种结论的依据是Pn与气孔导度以及它们与主要生态因子的相关性, 如张志华(1993)、路丙社(1999)、苗德全(1991)、陶俊(1999)等认为生态因子如水分亏缺、低空气湿度、强光和病害等原因导致气孔导度或气孔阻力的变化, 尽而形成了Pn “午休”和波动。少数也发现是由于非气孔因素的调节作用的结果(张大鹏, 1991; 黄丛林, 1996)。对喜树的研究结果表明气孔导度与Pn的相关性达到了显著水平, 午间喜树叶片的气孔导度也确有明显下降, 但随着气孔导度的降低, 午间CO2 int不但没有降低反而升高, 气孔的关闭没有减少对叶片CO2的供应, 说明“午休”的形成不是由于气孔导度下降所造成。从蒸腾强度的日变化为单峰曲线来看, 午间的蒸腾强度也没有降低, 进一步说明通道没有被关闭。由此可以认为喜树Pn“午休”的形成和波动是由非气孔因素造成的, 如叶肉阻力、叶肉的光合能力等, 具体因素还需进一步研究确定。
程来亮, 罗新书, 杨兴洪. 1992. 田间苹果叶片光合速率日变化的研究. 园艺学报, 19(2): 111-116. |
丁希泉, 郑秀梅编著.农业实用回归分析.吉林: 吉林科学技术出版社, 1989
|
冯建灿, 张玉洁, 谭运德, 等. 2000. 喜树与喜树碱开发利用进展. 林业科学, 36(5): 100-108. DOI:10.3321/j.issn:1001-7488.2000.05.017 |
郭志华, 王伯荪, 张宏达. 1998. 银杏的蒸腾特性及其对遮荫的响应. 植物学报, 40(6): 567-572. DOI:10.3321/j.issn:1672-9072.1998.06.014 |
黄丛林, 张大鹏. 1996. 葡萄光合速率日间降低内外因调控的研究. 园艺学报, 23(2): 128-132. |
郎奎建, 唐守正. 1989. IBMPC系列程序集. 北京: 中国林业出版社.
|
李建华, 罗国光. 1996. 巨峰葡萄叶片生长动态与光合特性的研究. 园艺学报, 23(3): 313-317. |
刘旭峰, 樊秀芳. 1993. 猕猴桃幼树光合特性的研究. 园艺学报, 20(4): 329-333. |
路丙社, 白志英, 董源, 等. 1999. 阿月浑子光合特性及其影响因子的研究. 园艺学报, 26(5): 287-290. DOI:10.3321/j.issn:0513-353X.1999.05.002 |
苗德全, 李淑美, 刘欣, 等. 1991. 田间茌梨叶片光合速率的日变化. 植物生理学通讯, 27(4): 273-275. |
彭永宏, 章文才. 1994. 猕猴桃的光合作用. 园艺学报, 21(2): 151-157. DOI:10.3321/j.issn:0513-353X.1994.02.019 |
沈允钢, 施教耐, 许大全. 1998. 动态光合作用. 北京: 科学出版社, 55-60.
|
陶俊, 陈鹏, 佘旭东. 1999. 银杏光合特性的研究. 园艺学报, 26(3): 157-160. DOI:10.3321/j.issn:0513-353X.1999.03.004 |
许大全. 1990. 光合作用"午睡"现象的生态、生理与生化. 植物生理学通讯, (6): 5-10. |
张大鹏, 黄丛林, 王学臣, 等. 1995. 葡萄叶片光合速率与量子效率日变化的研究及利用. 植物学报, 37(1): 23-25. |
张贤珍等编著. BASIC语言农业统计计算程序.北京: 农业出版社, 1990: 186~189
|
张志华, 高仪, 王文江, 等. 1993. 核桃光合特性的研究. 园艺学报, 20(4): 319-323. |
郑丕尧主编.作物生理学导论.北京: 北京农业大学出版社, 1992
|
朱林, 温秀云, 李文武. 1994. 中国野生种毛葡萄光合特性的研究. 园艺学报, 21(1): 31-34. DOI:10.3321/j.issn:0513-353X.1994.01.018 |
Farquhr G D, Sharkey T D. 1982. Stomatal conductance and photosynthesis. Ann. Rev. Plant Physiol, 33: 317. DOI:10.1146/annurev.pp.33.060182.001533 |
Hsiang Y, Liu L F. 1988. Identif ication of Mammalian DNA topoisomerase Ⅰ as an intracellular target of the anti-cancer drug camptothecin. Cancer Research, (48): 1722-1726. |
Liu Z, John Adams. 1996. Camptothecin yield and distribution within Camptotheca acuminata trees cultivated in Louisiana. Can. J. Bot., 74: 360-365. DOI:10.1139/b96-045 |
Priel E S, Showalter D, et al. 1991. Inhibition of human-immunodeficiency-vi rus (HIV-1)replication in vi tro by noncytotoxicdoses of camptothecin, a topoisomerase-Ⅰ inhibitor. AIDS Research and Human Retroviruses, 7(1): 65-72. DOI:10.1089/aid.1991.7.65 |
Priel E, Yosef O, et al. 1989. Detection of a novel DNA topoisomerase Ⅰ activity associatedwith human immunodeficiency virus (HIV)and other retrovirus particles. International Conference on Aids, 5: 586. |
Wall M E, Wani W C, Cook C E. 1966. Plant antitumor agent Ⅰ. The isolation and structure of camptothecin, a novel alkaloidal leukemia and tumor inhibitor from Camptotheca acuminata. J. Amer. Chem. Soc., 88: 3888-3890. DOI:10.1021/ja00968a057 |