 2002, Vol. 38
2002, Vol. 38文章信息
- 刘建军, 王得祥, 雷瑞德, 吴钦孝.
- Liu Jianjun, Wang Dexiang, Lei Ruide, Wu Qinxiao.
- 秦岭林区天然油松、锐齿栎林细根周转过程与能态变化
- TURNOVER PROCESS AND ENERGY CHANGE OF FINE ROOTS OF PINUS TABULAEFORMIS AND QUERCUS ALIENA VAR. ACUTESERRATA NATURAL FORESTS IN QINLING MOUNTAINS
- 林业科学, 2002, 38(4): 1-6.
- Scientia Silvae Sinicae, 2002, 38(4): 1-6.
-
文章历史
- 收稿日期:2001-07-31
-
作者相关文章


森林生态系统的结构、功能始终是生态系统研究的焦点。国内外最新研究结果表明, 细根是森林生态系统中最活跃的部分, 在森林生态系统的能量流动和物质循环两大生态过程中起着重要的作用(金明仕, 1992; 单建平等, 1992), 为了不断探讨生态系统中各分量的功能和作用, 对秦岭林区天然油松、锐齿栎林细根周转过程及能量变化规律进行了初步研究, 以丰富和完善森林生态系统的能量流动和物质循环领域的研究内容。
1 研究区自然概况研究区设在秦岭南坡火地塘林区, 位于北纬33°18′~33°28′, 东经108°20′~108°39′, 属长江水系的汇流区域。林区海拔高度变化于900~2450 m之间。区内山势陡峭, 地势破碎, 平均坡度在35°左右。
本林区处于北亚热带北缘, 年平均气温8~10 ℃, 年降雨量900~1200 mm, 年蒸发量800~950 mm, 湿润系数为1.022, 年总日照时数1100~1300 h, 无霜期199 d。
林区的主要成林树种有:油松(Pinus tabulaeformis Carr.), 锐齿栎(Quercus aliena var. acuteserrata Maxim.), 华山松(Pinus armandi Franch.), 红桦(Betula albo-sinensis Burk.), 光皮桦(Betula luminifera Winkl.), 青杄(Picea wilsonii Mast.), 冷杉(Abies fargesii Franch.), 山杨(Populus davidiana Dode.), 漆树(Toxicodendron vernicifluum (Stokes) F. A. Barkl.)。主要伴生树种有:青榨槭(Acer davidii Franch.), 千金榆(Carpinus cordata Bl.), 木姜子(Litsea pungens Hemsl.)等。土壤主要为花岗岩和变质花岗岩母质上发育起来的山地棕壤, 一般土层厚度约30~50 cm。
2 研究方法 2.1 林分调查在秦岭火地塘林区的中山地带选择立地条件相似的天然油松、锐齿栎林设置20m×20 m的标准地进行植被概况调查, 并进行每木检尺计算林分特征。油松林, 林龄60 a, 林分密度1950株·hm-2, 林分组成8油2栎, 平均胸径22.2 cm; 锐齿栎林, 林龄65 a, 林分密度1225株·hm-2, 林分树种组成8栎2漆, 平均胸径23.3 cm。
2.2 植物组织热值测定将样品烘干至恒重, 粉碎后置于干燥器中备用。采用GR-3500型氧弹式热量计测定根系热值(王得祥等, 1999), 每个样品测2~3个重复, 取两次误差不大于2 %的平均值作为测定结果。
热值计算公式:
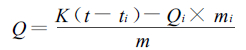
|
式中, Q为样品热值(J·g-1); K为热量计水当量(g); t为测定的主期温差(℃); ti为热量计热交换校正值(℃); Qi为引火丝燃烧热(J·g-1); mi为消耗的引火丝质量(g); m为样品质量(g)。
2.3 植物粗灰分含量采用干灰分法测定。
2.4 细根储存能量测定采用分层土柱结合水洗筛选法(W.伯姆, 1985)测定细根生物量, 根据热值换算为能量。在设定的油松、锐齿栎林标准地内随机挖取10个20 cm×20 cm×20 cm土柱, 带回实验室, 将土柱放入水中浸泡24 h, 清除砾石等杂物, 分拣出容易辨认的细根, 然后采用0.5mm的土壤筛冲洗土柱并分捡出所有细根; 根据根系的外形、颜色和弹性, 区分出≤5mm的活根和死根并称重。取一定量的根样在105 ℃条件下烘干至恒重, 计算干重, 根据热值将样品全部换算成能量。
2.5 细根储存能量动态测定及细根分解实验1997-04~10, 每月10~15日, 采用上述的土柱结合水洗筛选法测定细根(活根、死根)生物量, 并换算为能量。
用尼龙网袋法测定细根分解速率。4月初采集油松、锐齿栎活细根, 洗净风干后修剪成5 cm小段, 取样烘干测定含水率(以便换算成干重), 将风干的5 cm细根小段按树种装入20 cm×20 cm, 网眼为0.5 mm的尼龙网袋内, 每袋10 g左右, 用标签做好标记, 然后放入各自的样地内, 每个样地共投放了50袋。由于细根多分布在0~20 cm的土层内, 分解袋埋在离地表 10 cm的土壤中。每两个月在各样地内收取一次分解袋, 每个样地每次收取5袋, 用于失重分析和热值分析。
2.6 细根年生长积累能量、死亡归还能量和周转率计算根据McClaugherty等(1982)人提出的最大、最小值法, 利用改进的计算模型计算油松、锐齿栎细根的年生长积累能量、死亡归还能量和周转率。
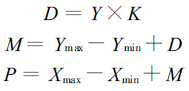
|
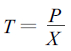
|
式中, D为细根年分解量; Y为死细根平均储存量; K为细根年分解率(由细根分解实验结果可以求得); M为细根年死亡损失量; Ymax为死细根储存量最大值; Ymin为死细根储存量最小值; P为细根年生长积累量; Xmax为活细根积累量最大值; Xmin为活细根积累量最小值; T为细根年周转率; X为活细根平均积累量。
3 结果与分析 3.1 细根分解过程能态变化林木细根的凋落、分解、归还构成了土壤库中能量转化和流动过程之一。有科学家指出, 在研究进入土壤中的植物残体及其腐殖化的问题时, 不应忘记具有理论和实际意义的腐殖质形成过程的能量问题(张一平等, 1988)。对油松、锐齿栎细根分解过程热值的测定结果表明(图 1):细根在分解过程中, 能态变化呈现波动起伏形式, 分解初期(油松60 d, 锐齿栎120 d)有一个吸能过程, 因此在这个时期, 单位质量的热值是增加的, 因为一些低能态的糖和脂肪, 被微生物迅速分解, 形成了高能态的微生物生物量。随着分解的继续, 高能态的有机物质逐渐被分解, 直到分解6个月后, 分解物的能态开始低于原始的细根, 即产生能量的损耗。这与张一平等(1988; 1996)研究植物残体分解过程中的能态特征相似。可见, 不能把分解能态变化简单地归结为放能或吸能过程, 而应被认为是既有吸能又有放能的一个错综复杂的能态变化过程。进一步分析油松、锐齿栎细根分解过程热值变化与灰分含量的关系, 灰分含量与热值呈负相关关系, 即灰分含量高, 分解物的热值低; 灰分含量低, 分解物的热值高。其回归关系式为:
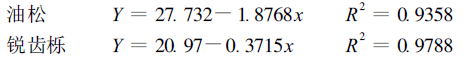
|

|
图 1 油松、锐齿栎细根分解过程的热值变化 Fig. 1 Caloric values variation of the fine roots during the decomposition for P. tabulaeformis and Q. aliena var. acuteserrata forests
 油松P. tabulaeformis; 油松P. tabulaeformis;  锐齿栎Q. aliena var. acuteserrata 锐齿栎Q. aliena var. acuteserrata
|
式中, Y为热值(kJ·g-1); x为灰分含量(%)。
3.2 细根分解过程及能量损失以上只是分析了油松、锐齿栎细根在分解过程中残留物的热值变化。事实上细根死亡凋落以后, 在微生物作用下逐渐分解矿化。表 1列出的是油松、锐齿栎细根分解能量损失变化, 可以看出, 分解时间越长, 各树种的能量残留率越小。特别是在开始的6个月分解速度较快, 残留率明显降低, 油松细根分解183 d的干物质残留率为65.1 %, 锐齿栎为61.1 %, 这是因为新近死亡的细根中含有大量的可溶性糖等容易被微生物分解利用的物质, 同时又恰逢水热条件最丰沛的时期, 所以这个时期分解较快。此后, 由于残留物中木素、纤维素等难以分解的物质比例增大, 分解速度逐渐减小。在经过1 a的分解后, 细根的能量残留率油松为46.8 %, 锐齿栎为42.9 %。大量的研究表明, 凋落物分解过程中的能量变化状况可以用衰减指数模型来描述:
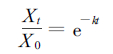
|
|
|

式中, X0为分解初始能量; t为分解时间(d); Xt为分解t d后的残存能量; K为分解系数即分解速率; e为自然对数的底(2.71828)。
运用衰减指数模型对试验结果进行回归分析(表 2)表明, 锐齿栎的分解系数为0.0031, 分解50 %干物质所需要的时间为250 d, 分解95 %干物质所需要的时间为993 d; 油松分解系数为0.0024, 分解50 %干物质需要的时间为312 d, 分解95 %干物质需要的时间为1272 d。比较两树种可见, 锐齿栎林细根分解比油松林快, 这与锐齿栎根系灰分含量较高, 有利于微生物分解有关; 而油松细根则含有大量松脂、单宁等不易被微生物分解的物质。与其他地区的研究结果相比, 秦岭火地塘林区油松细根的分解速率高于长白山阔叶红松林的0.0017(单建平等, 1993), 但低于会同杉木林的0.0026(廖利平等, 1995); 锐齿栎细根分解速率低于会同火力楠的0.0072(廖利平等, 1995), 表明不同树种、不同气候区林木细根的分解速率差异较大。
|
|

细根在一年中处在不断的生长和代谢凋落过程中, 通过对秦岭林区天然油松、锐齿栎林细根储存能量的季节动态的调查(表 3)表明, 油松活细根全年平均储存能量为119.43×106 kJ·hm-2, 变化范围为108.89×106~129.09×106 kJ·hm-2, 最大相对变幅为16.9 %; 锐齿栎活细根全年平均储存能量为172.70 ×106 kJ·hm-2, 变化范围为159.81×106~184.71×106 kJ·hm-2, 最大相对变幅14.4 %。两树种一年中活细根储存能量变化幅度差异不大。油松死细根全年平均储存能量为18.88×106 kJ·hm-2, 变化范围为12.93×106~24.20×106 kJ·hm-2, 最大变幅60.5 %; 锐齿栎全年平均储存能量45.2×106 kJ·hm-2, 变化范围为30.45×106~59.33×106 kJ·hm-2, 最大变幅61.9 %。相比之下, 同一树种1 a中死细根变化幅度较大, 活细根变化幅度较小, 即死细根储存能量的相对变化幅度大于活细根, 这可能与不同季节死细根分解速率变异较大有关。
|
|
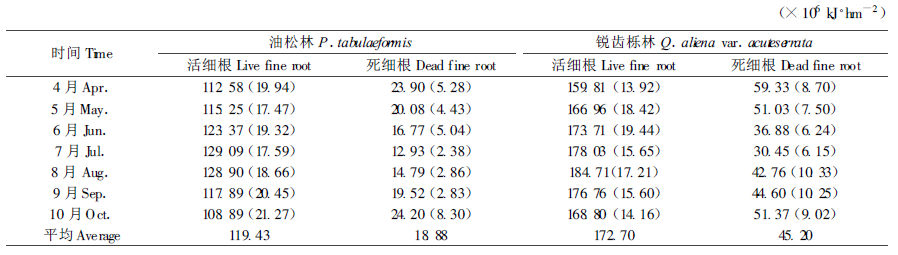
对不同季节细根储存能量的动态研究表明, 油松活细根储存能量以7月份最高, 生长季后期的10月份最低, 而死细根储存能量恰好相反, 以7月份最小, 10月份最大; 锐齿栎活细根储存能量则以8月份最高, 放叶之前的4月份最低, 而死细根储存能量则以4月份最大, 7月份最小。细根储存能量的这种变化可能与油松、锐齿栎在1 a中的生长节律有关。油松是常绿针叶树, 一般在生长季初期, 新生枝梢开始萌发前, 就有一定数量的碳积累, 为了满足这个时期生长对水分和养分的需求, 必须具有相应的适应机制, 以充足的细根参与养分吸收。特别是在7月份, 油松的材积生长最大, 必须以最多的吸收根参与水分和养分的吸收。到了9月份以后, 油松的材积生长已经非常缓慢, 这时一部分细根老化, 为了减少不必要的呼吸消耗, 细根开始大量凋落, 所以这个时期死细根储存能量明显增大, 直到生长季末的10月份林木开始休眠时达到最大。锐齿栎在5月初才开始放叶, 一般在6月末叶干重趋于稳定, 林分叶面积指数达到最大, 到8月份锐齿栎的材积生长最快, 需要的吸收根量也就最多。生长季后期, 树木生长减缓, 逐渐进入休眠期, 部分根系也如叶子一样凋落归还, 死亡量增大, 直到整个休眠期都有细根死亡, 因此翌年春季死细根储存能量达到最大。
3.4 油松、锐齿栎林细根年积累能量与周转研究天然油松、锐齿栎林细根的年死亡损失能量、生长积累能量及其周转率, 有利于阐明林木根系与土壤之间的能量交换。根据年分解率和死细根储存能量即可求得细根年分解释放量, 再运用Mc-Claugherty模型, 计算得出细根年死亡损失量、生长积累量及其周转率(表 4)。
|
|
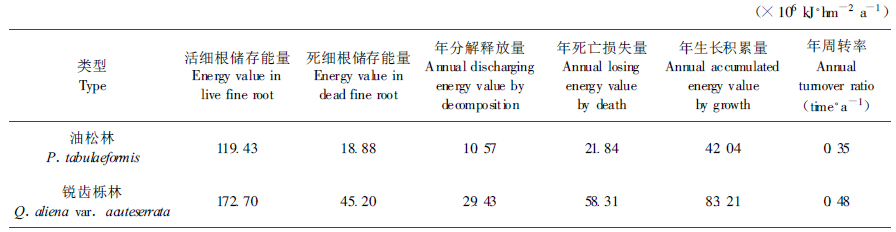
可以看出, 天然油松林活细根储存能量为119.43×106 kJ·hm-2 a-1, 只占乔木层生物总量的1.6 %, 锐齿栎林活细根储存能量为172.70×106 kJ·hm-2 a-1, 占乔木层生物总量的2.0 %, 细根在林分总储存能量中所占的比重较小, 这就导致人们在进行森林生态系统生产力研究时, 常常忽略了它的存在。但是油松林细根年死亡损失能量为21.84×106 kJ·hm-2 a-1, 占地上部分年凋落物损失能量的31.7 %, 锐齿栎林细根年死亡损失能量58.31×106 kJ·hm-2 a-1, 占地上部分年凋落物损失能量的62.8 %。考虑到细根分解速率比枯枝落叶高, 且养分含量也比枯枝落叶大(单建平等, 1993), 细根的死亡和分解成为林木归还土壤养分的重要途径, 因此进行森林生态系统养分循环和能量流动时, 必须充分重视林木细根的凋落与分解作用。
据有关报道, 细根的年周转率一般在0.29~1.20次·a-1之间。秦岭林区天然油松、锐齿栎林的细根周转率分别为0.35次·a-1和0.48次·a-1, 锐齿栎细根周转要比油松快。和空青山次生栎林的0.37次·a-1(陈金林等, 1999)相比也略高, 但与火力楠的1.42次·a-1(廖利平等, 1995)相比明显要低, 这与树种、林龄、土壤特性、气候条件以及人为干扰强度等因素有关。
4 结论细根的凋落、归还构成森林生态系统物质循环和能量流动的一个重要途径。基于能量研究细根的凋落和周转, 为开展不同生态系统中物质循环和能量流动研究提供了统一比较的基础。细根在分解过程中, 能态的变化不是一个简单的吸能或放能的过程。而是既有吸能又有放能的一个错综复杂的能态变化过程。能态的高低与分解物的灰分含量呈负相关。
细根分解过程符合指数衰减规律。锐齿栎的分解系数为0.0031, 年分解率为65.1 %; 油松分解系数为0.0024, 年分解率为56.0 %。
油松林活细根年均储存能量为119.43×106 kJ·hm-2, 变化范围为108.89×106~129.09×106 kJ· hm-2; 死细根年均储存能量为18.88×106 kJ·hm-2, 变化范围为12.93×106~24.20×106 kJ·hm-2。锐齿栎林活细根年均储存能量为172.70×106 kJ·hm-2, 变化范围为159.81×106~184.71×106 kJ·hm-2; 死细根年均储存能量为45.2×106 kJ·hm-2, 变化范围为30.45×106~59.33×106 kJ·hm-2。
油松林细根年周转率为0.35次·a-1, 年死亡损失能量为21.84×106 kJ·hm-2 a-1, 相当于地上部分年凋落物损失能量的31.7 %; 锐齿栎林细根年周转率为0.48次·a-1, 死亡损失能量58.31×106 kJ·hm-2 a-1, 相当于地上部分年凋落物损失能量的62.8 %。
陈金林, 许新健, 姜志林, 等. 1999. 空青山次生栎林细根周转. 南京林业大学学报, 23(1): 6-10. DOI:10.3969/j.issn.1000-2006.1999.01.002 |
[加拿大]J.P.金明仕.森林生态学.北京: 中国林业出版社, 1992
|
廖利平, 陈楚莹, 张家武, 等. 1995. 杉木火力楠纯林及混交林细根周转的研究. 应用生态学报, 6(1): 7-10. DOI:10.3321/j.issn:1001-9332.1995.01.005 |
单建平, 陶大立. 1992. 国外对树木细根的研究动态. 生态学杂志, 11(4): 46-49. |
单建平, 陶大立, 王淼, 等. 1993. 长白山阔叶红松林细根周转的研究. 应用生态学报, 4(3): 241-245. DOI:10.3321/j.issn:1001-9332.1993.03.008 |
王得祥, 雷瑞德, 尚廉斌, 等. 1999. 秦岭林区主要乔灌木种类能量背景值测定分析. 西北林学院学报, 14(1): 54-58. DOI:10.3969/j.issn.1001-7461.1999.01.011 |
W.伯姆. 1985. 根系研究法. 北京: 科学出版.
|
张一平, 白锦鳞, 等. 1988. 植物残体腐解过程中的能态变化特征. 西北农业大学学报, 16(4): 106-109. DOI:10.3321/j.issn:1671-9387.1988.04.020 |
张春慧, 张一平, 窦连彬. 1996. 农田生态条件下植物残体腐解过程腐解物的能态变化特征. 生态学报, 16(2): 140-146. |
McClaugherty C A, Aber J D, et al. 1982. The role of fine roots in the organic matter and nitrogen budgets of two forested ecosystems. Ecology, 63: 1481-1490. DOI:10.2307/1938874 |
Nadelhoffer K J, Aber J D, Melillo JM. 1985. Fine root s, net primary production, and soil nitrogen availability :a new hypothesis. Ecology, 66: 1377-1390. DOI:10.2307/1939190 |
Santantonio D, et al. 1987. Estimating fine root production and turnover from biomass and decomposition data. A compartment flow model. Can, J. For.Res, 17: 900-908. DOI:10.1139/x87-141 |