 2002, Vol. 38
2002, Vol. 38文章信息
- 雷相东, 唐守正.
- Lei Xiangdong, Tang Shouzheng.
- 林分结构多样性指标研究综述
- INDICATORS ON STRUCTURAL DIVERSITY WITHIN-STAND:A REVIEW
- 林业科学, 2002, 38(3): 140-146.
- Scientia Silvae Sinicae, 2002, 38(3): 140-146.
-
文章历史
- 收稿日期:2001-02-09
-
作者相关文章


生物多样性保护已成为森林可持续经营的一个重要目标。森林经营者需要了解森林经营活动后生物多样性的变化, 确保物种和种群不会因森林采伐和更新而处于危险境地, 从而对经营和保护做出正确决策。然而生物多样性被定义得太宽, 森林中的所有东西都被看成是生物多样性的象征(Silbaugh et al., 1997)。因此, 生物多样性指标的选择就很重要(Noss, 1999)。近年来在国际及国家层次上进行了生物多样性保护及指标的广泛合作, 我国也开展了生物多样性保护的研究(钱迎倩等, 1994; 蒋有绪等, 1998; 刘世荣等, 1998; 臧润国等, 1999)。生物多样性指标, 大致可分为3大类(Gustafsson, 2000) :物种和群落, 结构, 过程。作为一个很难定义好的集群概念, 物种只是生物多样性的一个不完整的替代指标(Bunnel et al., 1999)。林分结构是森林经营和分析中的一个重要因子, 是对林分发展过程如更新方式、竞争、自稀疏和经历的干扰活动的综合反映。林分中树木的空间布局决定了鸟类、昆虫、附生生物、下层植物及土壤微生物的生境的三维空间。近年来微观尺度上的生态学研究大部分认为空间上异质的森林能容纳更多的物种尤其是需要特殊生境的物种。如植被垂直结构的复杂性与一定面积中的昆虫和鸟类的数量有关(MacArthur et al., 1961)。多树种组成的林分, 径级或年龄结构变异越大, 林分结构越复杂, 能为各种动植物和微生物提供的小生境或食物的多样性就越大, 林分总体的生物多样性就越高(Buongiorno et al., 1994; Lahde et al., 1999a)。林分结构的变异性是林分潜在生物多样性的一个重要因素(Uuttera et al., 1995)。因此, 出现了一些基于结构的生物多样性指标。人们试图通过维持某种森林结构从而达到维持和增加生物多样性的目标。按是否考虑林分中树木的位置, 林分结构多样性指标可分为与距离有关的和与距离无关的两类。本文主要阐述这两类指标的研究进展及进一步要研究的问题。
1 与距离无关的林分结构多样性指标这类指标不考虑树木的位置。最常用的是反映树木的大小多样性。对于物种多样性指数, 当用直径、树高或其它结构分类变量来代替物种时, 它就可以反映林分结构多样性, 其中Shannon多样性指数最为常见(见公式1)。如Shannon树高多样性指数(Holland et al., 1994; Kuuluvainen et al., 1996)、叶层多样性指数(MacArthur et al., 1961)、垂直生境多样性指数(祝宁, 1982; 高宝嘉等, 1994)。
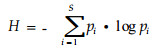
|
(1) |
式中, S为林分中结构分类变量的数量, pi为第i个分类变量的个体数占林分中总个体数的比。Shannon直径多样性指数或断面积多样性指数(Niese et al., 1992; Holland et al., 1994; Lin et al., 1996; Buongiorno et al., 1994, 1995; Volin et al., 1996)以及用联合熵来反映林分的树种、直径和树高的多样性1)。但是基于Shannon指数的林分结构多样性指标, 都需要将结构变量进行分组来计算相应的比例, 即需要将连续变量离散化, 如分成树高阶、径阶, 分组的方法和标准缺乏统一的依据。而分组的方法不同, 结构多样性结果也有差别。如研究林分垂直结构多样性时, MacArthur等(1961)将林分垂直结构分为0~0.61 m, 0.61~0.72 m, > 0.72 m或0~0.91 m, 0.91~9.14 m, > 9.14 m 3个层次; Kuuluvainen等(1996)则以2 m为树高级进行划分; Pretzsch (1999)则分为3层, 分别为林分最大树高的50%以下, 50%~80%, 80%以上。研究树木直径结构多样性时, Gove等(1995)采用5 cm径级分组; Lahde等(1999a)则分为3组(2~10 cm, 11~25 cm, > 25 cm)。因此, 这类分组主观性较强, 缺乏生态学意义。Latham等(1998)提出用树冠竞争点法对林分进行垂直分层, 具有一定的生态学意义。
雷向东.东北过伐林区几种典型森林类型的物种和林分结构多样性及采伐的影响研究.博士学位论文, 2000。
由于不同物种多样性指数比较结果的不一致性, Patil等(1982)提出了群落本质多样性(community intrinsic diversity)的概念, 用Δβ曲线或右尾和曲线来表示。它说明了生物多样性是一偏序集, 不是任何两个群落都可以进行多样性的比较, 存在二者在本质上不可比的情况, 这是一个观念上的创新。它克服了以往常用多样性指数比较结果的不一致性的缺点, 是一种与指数无关的多样性比较方法, 即如果两个群落存在本质多样性排序, 则按不同的多样性指数的排序结果应一致。只有本质多样性高的群落才具有真正高的多样性。据此, Gove等(1995; 1998)、Solomn等(1999)分别用右尾和曲线来度量树木的直径和树高结构多样性。对于不同林分间结构多样性的比较, 它是一种非常有用的方法。
另外, 基于林分中与物种多样性有关的一些结构成份, 一些学者建立了以结构为基础的林分多样性指数。如Pukkala (1997)用层次法提出的林分多样性指数包括以下的林分变量:枯死木蓄积(枯倒木和枯立木)、阔叶树蓄积、林分年龄、林分内的变异(物种混交、树木大小和林分密度等)。决策者可以赋予它们不同的权重。Lahde等(1999b)提出的林分内的多样性指数(INDFS)包括林分内的活立木、枯立木、枯倒木、分解木和特殊木(指特别老或大的树木, 珍稀树种), 表示为:

|
(2) |
式中INDFS为全林分的多样性指数, INDLT为活立木的多样性指数, INDDST为枯立木的多样性指数, INDDFT为枯倒木的多样性指数, INDCW为分解木的多样性指数, INDSP为特殊木的多样性指数并给出了前3种指数的非线性函数形式。实际上, 它们为断面积、株数及直径分布的函数。在野外调查时, 这个指数可做成一个评分表; 在研究和计算时, 可作为公式应用。它能较好地反映林分的发展阶段和立地类型。但他们均没有考虑林分的垂直结构。
以上这些指标虽然考虑了林分结构的不同方面, 但都没有直接把物种的水平和空间布局等空间特征考虑进去, 因此不能全面和细微地反映林分间结构多样性的差异。
实际上, 空间结构对于描述林分及其状态的改变有特别重要的意义。空间结构决定了林分内的生境和物种多样性, 对林分的发展有决定性的影响(Pretzsch, 1997)。因此, 出现了一些与距离有关的结构多样性指标。
2 与距离有关的林分结构多样性指标 2.1 反映树木的水平分布格局树木位置的水平分布格局, 可以分为随机、均匀和集群3种。确定分布格局的方法大致可分为样方法、距离法和二阶法(Kuuluvainen et al., 1996)。其中距离法中最常用的是最近邻体法(nearest neighbor analyses), 表 1列出了几种分布格局指数。
|
|

其中, 聚集指数R计算简单, 在林业上应用最广。另外, 惠刚盈等(1999a)提出的角尺度的概念, 通过描述相邻树木围绕参照树的均匀性来反映林分中林木的个体分布格局。与其它指数相比, 它的主要优点是不用测距离, 采用样线法即仅需调查样线上或距样线最近的树的角尺度就可获得分布信息, 从而得到林木在地域上的分布格局, 其结果的体现既可以用单个的角尺度值的分布也可用平均值。借助均值可将林分的林木个体分布评判为均匀、随机或集群, 但它不能准确地说明均匀、随机或集群的程度。
由于最近邻体法仅仅考虑了给定树木与其最近相邻木间的距离, 它受到小尺度格局的制约。二阶法中, 多用Ripley K函数等工具来研究林分中树木的二维水平点格局(Moeur, 1993; Kuuluvainen et al., 1996; 张金屯等, 1998)。由于Ripley的K函数分析考虑了所有树木间的距离, 可以分析各种尺度下的树木格局, 因此比最近邻体法更有效。Ripley的K函数由Ripley (1977)首先提出, 经Diggle (1983)等人发展, 其定义为:如果λ为单位面积的树木株数, 则λK (r)表示从一点出发距离为r的范围内期望的树木株数。K函数的形式为(Osher et al., 1981) :

|
(3) |

|
(4) |
式中, A为样地面积; n为树木株数; dij为树i与树j间的距离, 当dij < d时, I=1;dij≥d时, I=0;w为转化的边界修正, a、b为矩形样地的边长(a < b)。
对于矩形样地在最短边距离内对K函数的估计是无偏的, 它使计算更加简单并能在不同的尺度下进行分析(Penttinen et al., 1992)。为使其线性化和稳定方差, 将式(3)转化为:
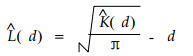
|
(5) |
在完全随机时, K函数的形式为:

|
(6) |
因此, 在完全随机分布时, 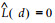
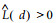
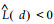
近年来, 分形理论也被用于植物种群的格局分析。马克明等(1999;2000a; 2000b)分别采用计盒维数、信息维数和关联维数对兴安落叶松种群进行了研究, 发现该种群在空间占据、个体分布复杂性和个体空间关联方面存在不同的尺度变化和分形特征, 将3者结合对该种群格局的综合分析, 能够得到较单一分形维数更全面、更准确的描述。
2.2 反映树种间的空间隔离关系Pielou (1961)根据最近邻体法, 用分隔指数S反映两个树种的空间隔离程度。其定义基于观察到的混交对和期望的混交对的关系。它能说明物种间的相互关系, S > 0时表示出现空间分隔; S=0表示两个物种相互独立; S < 0表示物种间相互吸引。计算分隔指数S需要的变量包括树种1和树种2的株数(m, n), 同种的相邻最近木的株数(a, d), 不同种的相邻最近木的株数(c, b), N为样地中树木的株数。从2×2列表(表 2)计算分隔指数S的公式为:
|
|

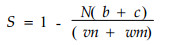
|
(7) |
但分隔指数仍是基于最近一株相邻木, 仅适用于分析树种随机分布混交林的种间隔离关系, 对均匀与团状分布的种群易造成不合理的描述。因此, 冯佳多等(1998)提出了单株混交度的概念, 用相邻最近单株中非同种单株所占的比例表示, 第i个单株的混交度Mi定义为相邻最近n株树木中非同种树种单株所占的比例,
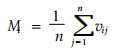
|
(8) |
vij为二元变量, 当第i个单株周围的第j个单株与i为同一树种时, vij=0;否则vij=1。惠刚盈等(2001)的研究表明, n=4能明显地表达出混交度的林学意义。此时, 混交度可分为5级:Mi=1为极强度混交, Mi=0.75为强度混交, Mi=0.50为中度混交, Mi=0.25为极弱度混交, Mi=0为零度混交。混交度表明了任意一株树的最近相邻木为其它种的概率。林分的树种混交度为林分中所有单株混交度Mi的平均值, 表示林分树种的混交程度。对于两个林分结构的比较, 可以用混交度各级别分布来直观反映, 但不能进行差异显著性检验。
以上提到的指标和方法都需要树木间的距离。他们没有考虑树木在空间的大小变化, 忽略了相邻木的大小和高度差异(Kuuluvainen et al., 1996), 而结构在任何时候都是具有水平和垂直成份的三维现象(Zenner, 2000)。
2.3 反映林分空间结构复杂性Kuuluvainen等(1996)用地统计学的方法研究经营林和原始林的空间结构多样性的差异。用半方差图(semivariogram)来说明林分中树木大小的空间自相关性和结构镶嵌体的尺度。半方差(semivariance)是一种反映样本之间空间相关性的指标。半方差γ (h)的定义为区域变量Z (x)在点x与点x+h处的值Z (x)与Z (x+h)差的方差的一半(王政权, 1999) :
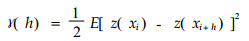
|
(9) |
其估计值为:
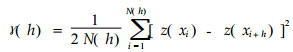
|
(10) |
式中, z (xi)、z (xi+h)为被距离h分开的两个位置的变量值; N (h)为被距离h分开的成对点的数目。随着点间距离的增加, 变量间差异增大, 半方差也随之增加, 直到点间距离大至它们彼此不相关。半方差能反映出统计意义上某个变量在不同尺度上的异质性, 如林分内不同大小树木的相互独立性。Kuuluvainen等(1996)用此分别研究了原始林和经营林分的胸径和树高的自相关性, 表明原始林的空间大小格局与经营林明显不同, 原始林的树木间空间自相关性较弱, 因此, 空间结构更为复杂多样。
但是, 半方差图能说明格局发生的尺度范围, 而没有直接提供结构复杂性方面的信息(Zenner et al., 2000)。一些学者基于参照木与n株相邻木间的大小关系来反映林分的空间结构复杂性。冯佳多等(1998)用大小分化度Ti来度量相邻树木的分化程度, 对于某个特定的单株树木i, 其Ti定义为:

|
(11) |
j=1, …n, n为相邻木的株数, di、dj分别为第i株和j株木的胸径(树高)。Ti值随相邻单株间平均大小分化程度的增加而增加, 分化度为零说明相邻单株大小相同。由于Ti均值的易混淆性和在复制结构时的不确定性等, 惠刚盈等(1999b)提出大小比数Ui的概念, 用大于参照树的相邻木占所考察的全部相邻木的比例表示:
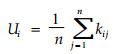
|
(12) |
kij为二元变量, 若相邻木j比参照木i小, kij=0;否则kij=1。其中n为相邻木的数量。
Ui值愈低, 说明比参照木大的相邻木愈少。它量化了参照木与相邻木的关系, 是对直径分布和大小分化度的完善和补充。与混交度类似, 林分的大小比为林分中所有单株大小比的平均值, 表示林分的分化程度。对于两个林分结构的比较, 可以用相应的分布来直观反映, 但也不能进行差异显著性检验。
以上指标没有把林分的三维结构综合为一个指数, 在进行林分结构多样性的比较时, 比较困难。
Zenner等(2000)基于树木位置的空间镶嵌(spatial tessellation)概念, 把水平点格局转化成三角面。认为每株树都看成是三维空间(x, y, z)中的一个不规则的空间数据点, 其中x、y为每株树的水平位置坐标, z为每株树的其它属性如树高或胸径。这样, 3株相邻木在三维空间中组成一个三角面, 整个林分组成一个镶嵌状的不重叠的不规则三角网(triangulated irregular network)。由此提出了一个结构复杂性指数SCI, 其定义为林分中所有不规则三角网的表面积和(SCI*)与其投影面积和(AT)之比。
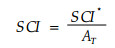
|
(13) |
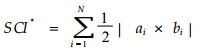
|
(14) |
式中, i=1, …N, N为三角网的个数; |ai×bi|为向量AB和向量AC的积, AB的坐标为ai= (xib-xia, yib-yia, zib-zia), AC的坐标为bi= (xic-xia, yic-yia, zic-zia)。
显然, 当所有树木具有同样树高时, SCI有最小值1。它的优点是把林分的垂直成份和水平成份结合起来。与Shannon树高多样性指数相比, SCI能更显著地反映林分的结构异质性差异, 能较灵敏地反映相邻木的差异和不同分布格局间的差异。如集群或随机分布的SCI值要比均匀分布的SCI值高, 但垂直分布对SCI的影响比水平空间格局分布更大。
另外, 借用景观生态学中斑块的概念, Zenner等(2000)把“斑块”定义为树高差小于一给定值的相邻树木的集合, 斑块类型定义为处于树冠层中一定高度层的斑块, 垂直梯度定义为组成三角形的3株相邻树木的最大树高差。这样, 林分就可以看成由斑块组成的结构镶嵌体。据此, 提出了另外几个结构复杂性指标如平均垂直梯度、斑块类型的平均数量、垂直梯度多样性指数和斑块类型多样性指数。它们与传统的结构变量如林分密度有强相关。在描述林分水平和垂直结构的复杂性时互相补充, 把林分的垂直结构复杂性定义为单位面积的斑块类型的数目。利用这些概念, 景观生态学的一些景观多样性指标和指数都可以用来进行林分结构多样性的测度。实际上, 其思想仍是以相邻木之间的关系为基础, 但利用了三维空间, 已扩展到面的概念, 给出了一个复合指标。但斑块的确定仍具有主观性。
综上所述, 林分结构多样性指标仍是一个研究热点。与距离无关的林分结构多样性指标没有考虑树木的空间位置, 对于整个林分大部分只能提供一个单一的平均值, 在进行不同林分间的比较时有一定的局限; 而与距离有关的林分结构多样性指标则考虑了树木的相对位置, 能更好地反映林分间结构的细微差异, 但由于需要距离或相对位置也限制了它的应用。因此, 林分尺度的结构多样性指标差别较大, 已经从一维发展到三维, 反映林分结构的各个方面。
3 问题与讨论 3.1 结构多样性指标的优点与物种多样性指标相比, 结构多样性指标作为生物多样性的一种替代, 容易测定, 调查费用相对较低, 容易纳入经营规划, 且具有可操作性。林分结构直接受到森林经营的影响, 因此, 基于林分结构的多样性指数容易同林分因子如断面积、密度和径阶分布等与经营直接有关的因子联系起来, 从而可以通过营林活动来实现理想的维持生物多样性的林分结构。因此, 它尤其适用于用材林的生物多样性保护。一些结构多样性指标已同生长模型相结合用于森林可持续经营规划和决策(Buongiorno et al., 1994; 1995; Volin et al., 1996; Gove, 1998)。
3.2 需要解决的问题还没有一个结构多样性指标能包括林分结构的所有成份, 因此, 指标的构建仍需进一步研究。结构不仅仅包括物种在空间和时间上的多度, 而且包括资源分配、生态位关系、个体大小关系、年龄分布、垂直、水平分布等。要既能体现林分结构的多维性, 又能灵敏地反映不同林分的结构差异, 且能通过模型与经营活动相联系, 利用模型来模拟和预测结构多样性, 通过经营措施来改变林分结构, 从而维持和增加生物多样性。另外, 一些结构多样性指标的抽样调查方法仍需要进行研究。
结构变量作为物种多样性的替代, 前提是越复杂的生境会支持物种更大的变异性(MacArthur et al., 1961), 且已得到经验性观测的支持。但也有例外(Hansson, 1994)。目前, 大多数这方面的研究都在局部范围, 一个地区的研究结论从多大程度上可以应用到另一地区还不清楚。但是, 已有大量的关于生物多样性同一定的结构和过程变量间关系的科学研究, 需要把这类研究进行集成, 把结果明确给林业工作者。因此, 仍需要检验林分结构多样性指标值, 研究各森林类型、不同地域林分结构多样性同物种多样性及相应的生态过程间的关系, 尤其是长期关系需要进一步研究和验证。
应把与结构有关的变量纳入现存的森林调查和规划体系。在林分层次如树种组成(包括幼树的密度、断面积和平均胸径)、枯立木的密度、可以区分的各林层的高度、灌木层的平均高等都可以纳入现存的森林调查规划体系。
高宝嘉, 张执中, 李镇宇.封山育林对植物群落结构及多样性的影响, 见: 徐化成, 郑均宝主编, 封山育林研究.北京: 中国林业出版社, 1994: 124~131
|
惠刚盈, Gadow K V, Albert M. 1999a. 角尺度——一个描述个体分布格局的结构参数. 林业科学, 35(1): 37-42. |
惠刚盈, Gadow K V, Albert M. 1999b. 一个新的林分空间结构参数——大小比数. 林业科学研究, 12(1): 1-6. |
惠刚盈, 胡艳波. 2001. 混交林树种空间隔离程度表达方式的研究. 林业科学研究, 14(1): 23-27. DOI:10.3321/j.issn:1001-1498.2001.01.004 |
蒋有绪, 郭泉水, 马娟. 1998. 中国森林群落分类及其群落学特征. 北京: 科学出版社, 中国林业出版社, 307-345.
|
克劳斯·冯佳多, 惠刚盈著.森林生长与干扰模拟.IUFRO-SPDC Textbook Project No.2.1998, 118~135
|
刘世荣, 蒋有绪, 史作民等著.中国暖温带森林的生物多样性.北京: 中国科学技术出版社, 1998, 1~195
|
钱迎倩, 马克平. 1994. 生物多样性研究的原理与方法. 北京: 中国科学技术出版社, 1-237.
|
马克明, 祖元刚, 倪红伟. 1999. 兴安落叶松种群格局的分形特征——关联维数. 生态学报, 19(3): 353-358. |
马克明, 祖元刚. 2000a. 兴安落叶松种群格局的分形特征——计盒维数. 植物研究, 20(1): 104-111. |
马克明, 祖元刚. 2000b. 兴安落叶松种群格局的分形特征——分形维数. 生态学报, 20(2): 187-192. |
臧润国, 刘静艳, 董大方著.林窗动态与森林生物多样性, 北京: 中国林业出版社, 1999, 184~221
|
张金屯. 1998. 植物种群空间分布的点格局分析. 植物生态学报, 22(4): 344-349. DOI:10.3321/j.issn:1005-264X.1998.04.008 |
祝宁. 1982. 生境及其多样性的测定. 东北林学院学报, (1): 134-141. |
王政权. 1999. 地统计学及在生态学中的应用. 北京: 科学出版社, 1-193.
|
Blackman G E. 1942. Statistical and ecological studies in the distribution of species in plant communities.I.Dispersion as a factor in the study of changes in plant populations. Ann.Bot, 6: 351-370. DOI:10.1093/oxfordjournals.aob.a088411 |
Bunnell F, Huggard D. 1999. Biodiversity across spatial and temporal scales:problems and opportunities. For.Ecol.Manage, 115: 113-126. DOI:10.1016/S0378-1127(98)00392-2 |
Buongiorno J, Dahir S, Lu H, et al. 1994. Tree size diversity and economic returns in uneven-aged forest stand. For.Sci, 40(1): 83-103. |
Buongiorno J. 1995. Growth and management of mixed species, Uneven-aged forests in the French Jura:Implications for economic returns and tree diversity. For.Sci, 41(3): 397-429. |
Clark Ph J, Evans F C. 1954. Distance to nearest neighbor as a measure of spatial relationships in populations. Ecology, 35(4): 445-453. DOI:10.2307/1931034 |
Diggle P. 1983. Statistical analysis of spatial point patterns. London: Academic Press.
|
Gove J H, Patil G P, Taillie C. 1995. A mathematical programming model for maintain structural diversity in uneven-aged forest stands with implications to other formulations. Ecological Modelling, 79: 11-19. DOI:10.1016/0304-3800(94)00044-I |
Gove J H.Optimizing the management of uneven-aged spruce-fir stands while preserving structural diversity.Bachmann P et al.(eds.), Assessment of biodiversity for improved forest planning, European forest institute proceedings.1998, 18: 123~134
|
Gustafsson L.Indicators and assessment of biodiversity from a Swedish forestry perspective.Forestry Research Institute of Sweden.2000, Report No.1
|
Hansson L. 1994. Vertebrate distributions relative to clear-cut edges in aboreal forest landscape. Landscape Ecology, 9: 105-115. |
Holland D N, Lilieholm R J, Roberts D W, et al. 1994. Economic trade-offs of managing forests for timber production and vegetative diversity. Can.J.For.Res, 24: 1260-1265. DOI:10.1139/x94-165 |
Kuuluvainen T, Penttinen A, Leinonen K, et al. 1996. statistical opportunities for comparing stand structural heterogeneity in managed and primeval forests:An example from boreal spruce forest in southern Finland. Silva Fennica, 30(2-3): 315-328. |
Lahde E, Laiho O, Norokorpi Y. 1999a. Diversity-oriented silviculture in the boreal zone of Europe. For.Ecol.Manage, 118: 223-243. DOI:10.1016/S0378-1127(98)00504-0 |
Lahde E, Laiho O, Norokorpi Y, et al. 1999b. Stand structure as the basis of diversity index. For.Ecol.Manage, 115: 213-220. DOI:10.1016/S0378-1127(98)00400-9 |
Latham P A, Zuuring H R, Coble D W. 1998. A method for quantifying vertical forest structure. For.Ecol.Manage, 104: 157-170. DOI:10.1016/S0378-1127(97)00254-5 |
Lin C R, Buongiorno J, Vasievich M. 1996. A multi-species, density-dependent matrix growth model to predict tree diversity and income in northern hardwood. Ecol.Modelling, 91: 193-211. DOI:10.1016/0304-3800(95)00190-5 |
MacArthur R H, MacArthur J W. 1961. On bird species diversity. Ecology, 42: 594-598. DOI:10.2307/1932254 |
Moeur M. 1993. Characterizing spatial patterns of trees using stem-mapped data. For.Sci, 39(4): 756-775. |
Niese J N, Strong T F. 1992. Economic and tree diversity trade-offs in management northern hardwoods. Can.J.For.Res, 22: 1807-1813. DOI:10.1139/x92-236 |
Noss R F. 1999. Assessing and monitoring forest biodiversity:A suggested framework and indicators. For.Ecol.Manage, 115: 135-146. DOI:10.1016/S0378-1127(98)00394-6 |
Osher J, Stoyan D. 1981. On the second-order and orientation analysis of planar stationary point process. Biometr.J, 23(6): 253-333. |
Patil G P, Taillie C. 1982. Diversity as a concept and its measurement. Journal of American Statistical Association, 77(379): 548-561. DOI:10.1080/01621459.1982.10477845 |
Penttinen A, Stoyan D, Henttonen H M. 1992. Marked point process in forest statistics. For.Sci, 38(4): 806-824. |
Pielou E C. 1959. The use of point to plant distances in the study of the pattern of plant populations. J.Ecol, 47: 607-613. DOI:10.2307/2257293 |
Pielou E C. 1961. Segregation and symmetry in two-species populations as studied by nearest neighbor relations. J.Ecol, 49: 255-269. DOI:10.2307/2257260 |
Pretzsch H. 1997. Analysis and modeling of spatial stand structure:Methodological consideration based on mixed beech-larch stands in Lower Saxony. For.Ecol.Manage, 97: 237-253. DOI:10.1016/S0378-1127(97)00069-8 |
Pretzsch H.Structural diversity as a result of silvicultural operations.In: Olsthoorn A F M et al.edited, Management of mixed-species forest: Silviculture and Economics, 1999: 157~174
|
Pukkala T. 1997. Integrating forest-level and compartment-level indices of species diversity with numberical forest planning. Silva Fennica, 31(4): 417-429. |
Ripley B. 1977. Modelling spatial pattern. J.Roy.Statist.Soc, 39: 172-242. |
Silbaugh J M, Betters D R. 1997. Biodiversity values and measures applied to forest management. Journal of Sustainable Forestry, 5(1/2): 235-248. |
Solomon D S, Jeffrey H G. 1999. Monitoring diversity over time using a forest growth model based on ecological land classfications. For.Ecol.Manage, 114: 265-275. DOI:10.1016/S0378-1127(98)00358-2 |
Uuttera J, Maltamo M. 1995. Impact of regerneration methods on stand structure prior to first thinning:Comparative study North Karelia, Finland vs.Republic of Karelia, Russian Federation. Silva Fennica, 29(4): 267-285. |
Volin L, Buomgiorno J. 1996. Effect of alternative management regimes on forest stand structure, species composition and income:a model for the Italian Dolomites. For.Ecol.Mnage, 87: 107-125. DOI:10.1016/S0378-1127(96)03828-5 |
Zenner E K, Hibbs D E. 2000. A new method for modeling the heterogeneity of forest structure. For.Ecol.Manage, 129: 75-87. DOI:10.1016/S0378-1127(99)00140-1 |