 2002, Vol. 38
2002, Vol. 38文章信息
- 刘家冈, 张学培, 刘宁, 张立, 王本楠.
- Liu Jiagang, Zhang Xuepei, Liu Ning, Zhang Li, Wang Bennan.
- 羊草草原弃耕地恢复性演替过程的数学模型和计算机模拟
- A MATHEMATICAL MODEL AND A COMPUTER SIMULATION OF SUCCESSION PROCESS OF LEYMUS CHINENSIS MICROCOMMUNITIES IN ABANDONED FIELD
- 林业科学, 2002, 38(3): 63-66.
- Scientia Silvae Sinicae, 2002, 38(3): 63-66.
-
文章历史
- 收稿日期:2001-02-28
-
作者相关文章


2. 中国科学院环境与生态研究中心 北京 100085
2. Research Center for Eco-environment Sciences, Chinese Academy of Sciences Beijing 100085
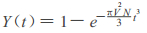 Where Y(t)is area occupied by microcommunities of Leymus chinensis, V the expansion speed of Leymus chinensis microcommunities, N the number of seed which invade the abandoned field in unit time and unit area.
Where Y(t)is area occupied by microcommunities of Leymus chinensis, V the expansion speed of Leymus chinensis microcommunities, N the number of seed which invade the abandoned field in unit time and unit area.防止荒漠化是我国环境保护的重要任务, 退耕还草则是防止荒漠化的基本方法之一, 恢复性演替又是退耕还草的一个主要技术途径。研究恢复性演替的规律, 显然具有重要的生态学意义和环境保护意义。本文根据王振堂等(1985)的研究资料, 提出我国东北地区弃耕地羊草恢复性演替过程的数学模型和计算机模拟。
羊草(Leymus chinensis)是中国东北地区西部草原上优良牧草, 常形成占绝对优势的群落。草原的兴衰与羊草群落的演替有密切的关系。王振堂等(1985)曾经在弃耕地上观察到羊草的恢复性演替过程, 进行了系统地研究。根据他们的研究, 弃耕地上羊草群落的恢复大体分为3个阶段: (1)种子在自然力的作用下, 以随机分布的方式重新侵入弃耕地并形成幼苗。(2)种子形成幼苗后, 经过无性繁殖过程, 由根茎生成分蘖植株, 以一定的平均速率向四周扩散, 逐渐形成圆形的羊草小群落。图 1是演替3~5 a后在草层背景中镶嵌的圆形羊草小群落, 景象别致。(3)羊草小群落不断扩大, 逐步合并成较大的群落, 最终融合成大片的羊草群落。图 2是演替7 a后的羊草小群落的分布, 可以看到一些羊草小群落已经开始合并, 形成较大的群落。表 1是在与图 2同一地段测得的羊草小群落的半径分布。

|
图 1 羊草小群落(王振堂, 1985) Fig. 1 Leymus chinensis microcommunity(Wang Zhentang, 1985) |

|
图 2 演替7 a的羊草小群落分布(王振堂, 1985) Fig. 2 Distribution of micro communities after seven years succession (Wang Zhentang, 1985)  羊草群落Leymas chinensis microcommunity; 羊草群落Leymas chinensis microcommunity;  其它草类Other species. 其它草类Other species.
|

但根据我们的理解, 第(1)阶段的重新侵入过程是逐步进行的, 它与第(2)、(3)阶段的过程互相重叠。也就是说, 当先侵入的种子已经形成幼苗, 并开始向周围扩展以后, 仍有种子陆陆续续地侵入到弃耕地中来, 后来的种子也会形成幼苗和小群落。这就说明了为什么在同一块弃耕地中, 不同的羊草小群落有不同的半径。事实上, 不同大小的羊草小群落有不同的“年龄”。
1 数学模型根据王振堂(1985)的研究和我们的理解, 我们假定, 羊草种子的侵入时间是均匀分布的随机序列, 并且以随机的地点在弃耕地上形成幼苗。随后, 便以一个平均速率向周围扩展。因此, 弃耕地上存在一个随机分布的羊草小群落序列, 其半径与其种子侵入到现在的时间成正比, 也就是说, 与羊草小群落的年龄成正比。
设样地是边长为L的正方形, 面积为SL; 而S1为第1个小群落的面积, 当只有第1个小群落时, 样地中没有羊草的面积占总面积的比例为
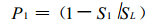
|
(1) |
同理, 当只有第2个小群落时, 样地中没有羊草的面积占总面积的比例为
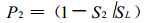
|
(2) |
由于两个小群落是随机分布的, 故当两个小群落同时存在时, 样地中没有羊草的面积占总面积的比例为
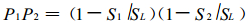
|
(3) |
同理, 当只有前n个小群落时, 样地中没有羊草的面积占总面积的比例为
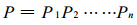
|
(4) |
其中,
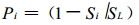
|
(5) |
因此, 此时样地中有羊草的面积为
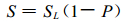
|
(6) |
让我们回过头来看看羊草小群落面积Si的计算,
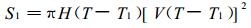
|
(7) |
式中, H (x)是Haivesid函数, 即当x > 0时, H (x) =1;当x < 0或x=0时, H (x) =0.T1是第1个羊草小群落发生的时间。Haivesid函数的作用在这里是保证羊草小群落在发生之前是没有面积的。余此类推。
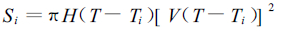
|
(8) |
Ti是第i个羊草小群落的发生时间。
根据(4)、(5)、(6)和(8)式可以计算羊草面积S。
虽然王振堂(1985)指出, 在早期(3 ~ 5 a), 羊草小群落内的植株密度很低, 要等到6 a以后, 密度才逐渐达到最大值。但为了模型的简洁, 我们忽略群落中羊草密度的变化, 而直接采用最大值。之所以这样做, 是因为处在早期的羊草小群落面积小, 个数少(它们很快就发展成为中、晚期小群落), 对整体的重要性相对较小的缘故。根据模型方法论, 模型与数据符合与否, 是判断在建模过程中忽略某些因素是否合理的重要客观依据。从后面可以知道, 我们模型的计算结果与数据符合得很好。
2 参数的估算根据表 1可知, 7 a大约发生了16个羊草小群落, 大约每年平均发生2.3个小群落, 即每0.44 a发生1个小群落。因此, Tn可看作是一个以0.44 a为公差的等差序列{ti}。虽然, 从草本群落的生物学特性看, 似乎不应该0.44 a发生一个小群落, 而应该取一个整数。但是我们建立的是连续模型, 0.44 a在这里只是一个时间参量、平均量而已。
根据表 1中第5组数据, 可以计算出羊草小群落向外扩展的平均速度大约为V=0.6 m·a-1。这与王振堂(1985)的计算不同, 王振堂(1985)将表 1中的16组数据当作是有相同年龄的羊草小群落的半径而加以平均。我们则认为它们应该属于一个年龄序列, 不同的半径对应不同的年龄。该样地是7 a演替弃耕地, 故只有最大羊草小群落(第5组)的年龄才应该为7 a。
根据王振堂(1985)的图 2 (或本文图 3)所示羊草植株密度的实测数据, 可推知7 a演替时样地的羊草植株的平均密度约为33.3棵·m-2。而根据表 1则可知7 a演替样地中羊草小群落总面积为333 m2, 占样地总面积(SL=80 m×80 m)的5.2%.因此可用33.3棵·m-2除以5.2%, 得小群落内羊草平均密度为W=640棵·m-2。由SW/SL可得样地羊草植株密度的计算值。

|
图 3 羊草小群落的扩展过程 Fig. 3 Expansion process of microcommunities ○王振堂(1985)的实测数据(0~20 a), Data (0~20 years) (Wang Zhentang, 1985); ——本文数学模型的计算结果, Result of the mathematical model; ○本文计算机模拟的结果, Result of the computer simulation; ·本文后面解析模型的计算结果, Result of analytic model. |
由(6)式计算的S值再乘以(W/SL), 便得到演替50 a全过程的计算结果(见图 3), 此结果与王振堂(1985)的数据十分符合。
3 计算机模拟按我们对羊草小群落恢复性演替的三个阶段的理解, 去实行计算机模拟。
在80 m×80 m的方块中, 每0.44 a随机地发生1个羊草小群落, 小群落一旦发生, 便以V=0.6 m·a-1的平均速率向外扩大。我们测量每1 a的羊草小群落的总面积(减去互相重叠的部分), 乘以羊草小群落内的羊草平均密度, 便得到当年羊草总棵数, 再除以SL求得到样方内羊草的平均密度。图 3中大圆圈表示计算机模拟的50 a的结果。这个结果与(6)式计算的结果十分一致, 也与王振堂(1985)的实测数据符合得很好。
从图 4可见, 已经发生的16个小群落, 形成一个序列, 随机地分布在样地上, 其特征在定性上与图 2的实际结果一致。
4 数学模型的解析形式根据(4)、(5)、(6)、(8)式, 有

|
(9) |

|
图 4 7 a演替羊草小群落的计算机模拟图 Fig. 4 Distribution of microcommunities of Leymus chinensis by computer simulation after seven-years succession 图中有16个小群落, 最大的7个圈, 代表 7 a的小群落, 最小的才1圈, 代表小群落刚刚发生。这些小群落有的已经互相融合(但仍然可分辨)。 There are 16 microcommunities. The largest one has 7 circles, which represents 7 years old microcommunity. The most little one only has 1 circle that means the microcommunity is just born. Some microcommunities have joined each other (but are still distinguishable). |
当样地面积SL→∞时, 根据近似公式, 有

|
(10) |
其中N是单位时间、单位面积内侵入的种子数。Δt是前后相继侵入的两粒种子的时间间隔。因为Δt=1/NSL, 所以(NSLΔt) =1, 乘以1是一个恒等变换。因为SL→∞时, 羊草种子侵入的时间序列{ti}变成Δt→0的无穷等差序列。则(10)式Z中的求和变为定积分:

|
即
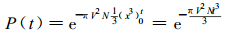
|
(11) |
则根据(6)式, 草地上有羊草的概率为
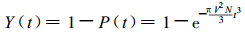
|
(12) |
(12) 式就是数学模型的解析形式。解析形式实际上是基本数学模型在样地趋于无穷大时的极限。
代入前边得到的参数, V=0.6 m·a-1, N=2.3个·a-1在80 m×80 m样方内, 所以,

|
(13) |
计算结果在图 3里, 与数据和其他模拟结果符合得很好。
5 结论与讨论对弃耕地内羊草小群落发生发展的研究, 不仅解释了羊草小群落恢复早期的形成机制, 解释了演替中期羊草小群落开始互相融合时的情况, 也解释了羊草群落最后融合成大片草原的过程。
实际上, 在羊草恢复性演替的早期(图 3的0~15 a), 羊草小群落面积小、数量少, 因而互相重叠少, 羊草的面积基本上是各个圆形羊草小群落面积的总和, 所以图 3中的羊草密度是以二次曲线形式增长。在演替的中期(图 3的15~25 a), 羊草小群落面积逐渐加大、数量增多, 它们就开始融合, 由于互相重叠的部分增多, 羊草密度逐渐由原来的二次曲线变得平直。当进入演替晚期时, 羊草群落大面积融合, 只剩下一些小片无羊草地块, 它们随着时间的推移而缩小、而消失, 因此羊草密度由直线进一步变成反二次曲线(图 3的25 a以后), 从而进入了羊草草原的顶极阶段。
(4)、(5)、(6)和(8)式, 是本文提出的计算羊草小群落恢复性演替的数学模型。其中(4)、(5)式与植被辐射理论中(刘家冈等, 1985a; 1985b)的透过函数的形式很相近。事实上, 本文作者正是觉得羊草小群落的随机分布和互相重叠, 与树叶在地面上的投影分布与重叠有某种相似性, 才悟出了羊草小群落的恢复性演替数学模型。这是概念和理论的创造技法中的“形象相似法”的应用, 也是创造性思维中形象思维重要性的体现。
王振堂, 祝廷成. 1985. 羊草小群落扩散过程的初步分析. 生态学报, 5(3): 213-222. |
复旦大学编. 1979. 概率论(第二分册, 数理统计). 北京: 人民教育出版社, 264-272, 401.
|
刘家冈, 张文杰. 1985. 光在林冠中的传播Ⅰ. 林业科学, 21(3): 234-240. |
刘家冈, 张文杰. 1985. 光在林冠中的传播Ⅱ. 林业科学, 21(4): 360-368. |