 2002, Vol. 38
2002, Vol. 38文章信息
- 李淑娴, 尹佟明, 邹惠渝, 丁雨龙, 黄敏仁.
- Li Shuxian, Yin Tongming, Zou Huiyu, Ding Yulong, Huang Minren.
- 用水稻微卫星引物进行竹子分子系统学研究初探
- PRELIMINARY STUDY ON MOLECULAR SYSTEMATICS OF BAMBOO BY SSR PRIMERS DERIVED FROM RICE
- 林业科学, 2002, 38(3): 42-48.
- Scientia Silvae Sinicae, 2002, 38(3): 42-48.
-
文章历史
- 收稿日期:2000-09-11
-
作者相关文章


竹类植物是禾本科的1个自然类群, 广泛分布于亚洲、拉丁美洲、非洲受季风气候影响、水热条件较好的热带、亚热带地区, 少数属种生长在温带乃至寒带地区。其水平分布从南半球拉丁美洲的阿根廷南部(47°S)到北半球的库页岛(46°N); 垂直分布从平原到高山雪线。竹类植物常常组成大面积竹林, 是森林资源的重要组成部分(温太辉, 1983)。
亚洲是世界竹类植物的分布中心, 无论属种数目及在生产上的重要性都较美洲、非洲为甚。而中国是世界上竹种资源最丰富的国家, 其属种远较世界各国为多。从黄河流域到海南岛, 从东海之滨直到西藏南部的24个省(区)都有竹子分布。因此, 中国竹子研究的进展对世界各国, 尤其是对亚洲地区的竹子研究具有很大的影响。
竹子研究中的1个突出问题表现在竹子的分类学研究上, 竹亚科的分类研究比其它种子植物困难, 主要原因是竹类很少开花, 开花的植株常不正常, 开花后又多死亡。而花是竹亚科分属及属以上分类等级的主要依据(耿伯介, 1982; 王正平等, 1983)。20世纪70年代以来, 中国竹子分类学研究取得了很大成就, 发现了不少具有重要经济和科学价值的竹子属种, 但同时, 在分类上也存在着严重的混乱和错误。竹亚科中散生竹的分布范围远较丛生竹为广, 其形态特征表现错综复杂, 属间界限连续不断, 难以划分(陈守良等, 1983), 因此散生竹的分类更为混乱。散生竹分类中最大的问题反映在青篱竹属分类的混乱上(杨光耀等, 1993)。青篱竹属(Arundinaria)是竹亚科散生竹类建立最早的属, 曾经是一个范围庞大、特征混杂的大属, 全世界先后发表为Arundinaria的双名近400个, 包括美洲、亚洲和非洲的很多竹种。随着竹子研究的深入, 美洲、非洲和亚洲南部的青篱竹属分类上的混乱已相继被澄清, 但东亚(中国和日本)的青篱竹属尚缺乏系统研究。一段时间以来, 竹子分类研究十分活跃, 新分类群不断发表, 而且相继建立了与青篱竹属关系十分密切的属, 如苦竹属(Pleioblastus), 矢竹属(Pseudosasa), 巴山木竹属(Bashania)等, 这些属与青篱竹属的归并, 有的是比较自然的, 有的却导致了更大的混乱, 以致东亚有无青篱竹属竹种分布成为东亚散生竹分类的争论焦点。现代分子生物学的发展为这一问题的研究提供了新的研究方法, 现代分子生物学方法与经典分类学研究方法的结合, 有望使这些问题得到解决。最有效的研究这些问题的方法是利用DNA测序技术, 根据同源基因家族的进化历程来推断各属间的系统进化关系。一般系统学研究中, 基因的变异要求在所研究的分类等级中具有5%~15%的变异(Olmsted et al., 1994; Steele et al., 1994; Li et al., 1997; 1999;Germano et al., 1999)。由于缺乏对青篱竹属相关属遗传背景的认识, 序列的选择上也有一定困难, 因此需要选择合适的研究手段首先对广义青篱竹属的各个相关属的亲缘关系进行研究。
根据竹子的分子生物学研究基础, 目前可供选择的主要为没有种属特异性、可通用的标记技术, 如RAPD和AFLP (amplified fragment length polymorphism)等。这些标记技术都不需要事先知道DNA序列的信息, 可用随机引物或人工合成的引物通过PCR反应获得。但如果所研究材料的亲缘关系较远, 等分子量的扩增谱带可能并不同源, 因此这些标记技术产生的谱带不适合用来研究较高分类等级物种的亲缘关系, 最适合的分类等级应为种下(Dario, 1991; 邹喻萍, 1995)。朱其慧, 尹佟明等(2000, 未发表)利用RAPD标记技术对青篱竹属相关属的亲缘关系进行了研究, 得到的结果较为混乱, 可能是这些竹子的亲缘关系相对较远, 不适宜用RAPD标记研究这些竹子的亲缘关系。Moore (1995)综合了禾本科不同物种的研究结果, 发现禾本科不同物种的基因组有相同的起源, 通过禾本科原始染色体片段的组合或重复, 可获得禾本科不同物种的基因组, 不同的基因组形成1个同心圆。而根据国际水稻所的研究结果, 竹子与水稻的亲缘关系是禾本科中较近的2个分支, 水稻的分子生物学研究基础是相当雄厚的, 有许多研究结果可直接应用到竹子的研究中来。其中利用水稻的RFLP探针来研究不同竹子间的亲缘关系就是1个非常有效的途径。另1方法是利用水稻的SSR引物来研究不同竹子间的亲缘关系, 虽然SSR引物有较高的种属特异性, 但在一定的分类等级上也具有相对的保守性(何平, 1998), 通过对水稻微卫星引物的筛选, 有望获得与竹子通用的微卫星引物。本研究利用中国科学院遗传所朱立煌研究员提供的94对水稻微卫星引物, 探讨了青篱竹属相关属竹种的亲缘关系。
1 材料材料采自南京林业大学竹类植物园, 于春季采集新竹上的初展叶片, 包括矢竹(Pseudosasa japonica矢竹属的模式种)、菲白竹(Arundinaria fortunei)、翠竹(Arundinaria pygmaea)、大明竹(Pleioblastus gramineus苦竹属的模式种)、斑苦竹(Pleioblastus maculatus)、苦竹(Pleioblastus amarus)、宜兴苦竹(Pleioblastus yixingensis)、茶秆竹(Arundinaria amabilis)、福建茶秆竹(Arundinaria convexa)、巴山木竹(Bashania fargesii巴山木竹属的模式种)。
2 方法 2.1 DNA提取DNA提取方法按照Couch等(1990)的方法稍加改进。将2~3片叶片放入研钵中并加入液氮研磨, 然后转入10 mL离心管中, 加入2 mL SDS DNA提取液[100 mmol·L-1 Tris (pH8.3), 5 mmol·L-1 EDTA (pH8.3), 500 mmol·L-1 NaCl, 1.5%SDS, 350 mmol·L-1 β-mercaptoethanol], 充分摇匀, 加入700 μL 5 mmol·L-1 KAc, 充分摇匀, 冰浴30 min, 8000×g离心10 min, 将上清液转入新的10 mL离心管中, 再加入3 mL酚:氯仿(1:1), 充分摇匀, 8000×g离心10 min, 将上清液转入新的10 mL离心管中, 加入3 mL氯仿, 摇匀, 8000×g离心10 min, 将水相转移至新的10 mL离心管中, 加入6 mL无水乙醇, 轻轻摇匀, 将离心管置于冰箱中沉淀30 min, 用广口吸管将浮起的成团沉淀物吸出, 转入1.5 mL离心管中, 10000×g离心10 min, 倒掉水相, 用1 mL 70%乙醇冼沉淀2次, 然后用无水乙醇冼沉淀1次, 将离心管倒置在吸水纸上, 于空气中干燥, 将沉淀物溶于100 μL TE中, 于-70℃存贮。DNA浓度用琼脂糖凝胶比色法与标准浓度的λDNA比较确定。
2.2 DNA扩增DNA扩增在PE-9600 PCR仪上进行, 反应体系(20 μL)组成如下:5 ng左右模板DNA, 正、反向引物各20 pmol, dATP, dCTP, dGTP, dTTP的量各为200 μmol·L-1, 1.5 μL 10倍的反应缓冲液[100 μmol·L-1 Tris-HCl (pH8.3), 500 mmol·L-1 KCl, 20 mmol·L-1 MgCl2, 0.01%明胶, 5.0 g·L-1 BSA, 1 mg·L-1 TMACl], 1U·μL-1 Taq DNA聚合酶(MBI产品)。扩增条件为:94℃预变性2 min, 接着进行94℃变性30 s, 55℃退火30 s, 72℃延伸1 min, 反应38个循环, 最后72℃链延伸10 min, 扩增产物用3%琼脂糖凝胶电泳检测分析。
2.3 数据收集及结果分析竹子与水稻的基因组存在一定程度的分化, 用水稻的SSR引物在竹子中扩增时会产生一些杂带, 为保证扩增位点的特异性, 在位点的选择上考虑到微卫星序列的长度相对较小, 同时竹子是多年生物种, 相应的微卫星序列长度可能比水稻基因组的长。在扩增过程中我们发现, 片段长度在400bp以下的谱带具有较好的重复性, 因此仅对长度小于400 bp的多态谱带进行了统计, 对获得的标记按显性标记处理。利用POPGENE32软件按照UPGMA法进行聚类分析。聚类过程中遗传距离用下式计算(Nei, 1983; Kimura, 1980) :

|
式中, Nxy为X、Y两个个体共享标记数; NX、NY、X、Y为个体所拥有的标记数; I为正规化位点共享度。
3 结果与讨论在所用的94对引物中, 有42对引物(44%)扩增出了强度不等的产物, 说明这些引物在竹子基因组中存在对应的同源序列。从中选出18对扩增产物较强的水稻SSR引物用于本研究, 这些引物的序列及在水稻基因组中的背景材料见表 1。这些微卫星序列分布在水稻基因组中除第4、第12染色体以外的不同染色体上, 在水稻基因组中分布相对比较均匀。
|
|
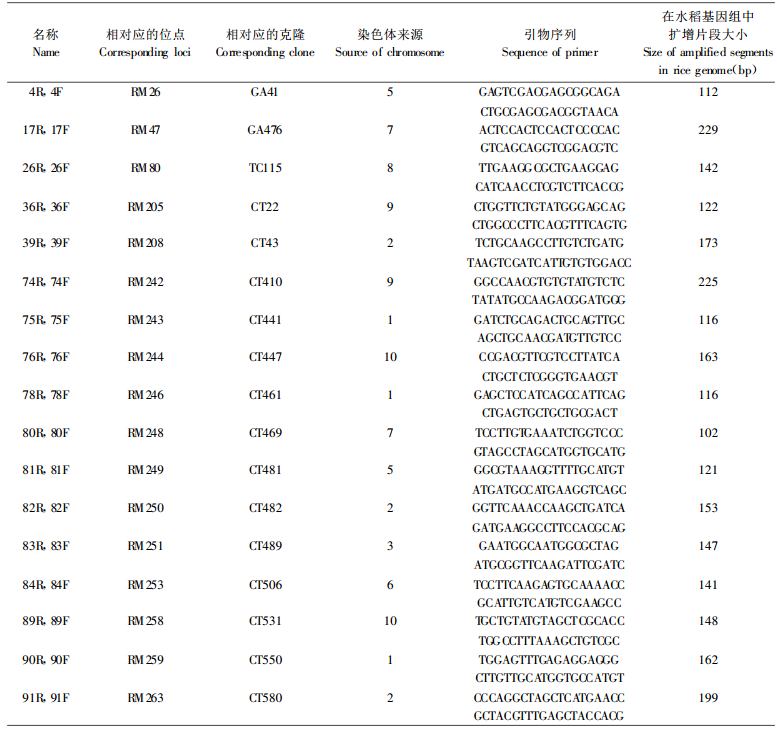
竹子和水稻在进化上存在较大的分化, 水稻的微卫星引物在竹子中很难有100%的同源性, 因此PCR反应过程中会产生一些杂带, 本研究对小于400 bp的扩增谱带进行了统计分析。所用的18对引物共产生了49个分子量小于400 bp的多态位点。图 1所示为引物组合81R, 81F及91R, 91F的扩增结果。

|
图 1 引物组合81R, 81F (a)及91R, 91F (b)对竹子DNA的扩增谱带 Fig. 1 The amplification results in the studied bamboo by primer combinations of 81R, 81F (a) and 91R, 91F (b) 1~10分别为矢竹(Pseudosasa japonica)、菲白竹(Arundinaria fortunei)、翠竹(Arundinaria pygmaea)、大明竹(Pleioblastus gramineus)、斑苦竹(Pleioblastu smaculatus)、苦竹(Pleioblastus amarus)、宜兴苦竹(Pleioblastus yixingensis)、茶秆竹(Arundinaria amabiliis)、福建茶秆竹(Arundinaria convexa)、巴山木竹(Bashania fargesii)。“-”为未加模板的对照。M为分子量标记。 No.1~10 are Pseudosasa japonica、Arundinaria fortunei、Arundinaria pygmaea、Pleioblastus gramineus、Pleioblastus maculatus、Pleioblastus amarus、Pleioblastus yixingensis、Arundinaria amabiliis、Arundinaria convexa、Bashania fargesii."-" is the contrastwithou tDNA.M i smolecularweigh tmarker. |
统计分析结果见图 2和表 2。可以看出, 属下分类等级的各个竹种的聚类结果与属下这几个种传统分类学的结论比较和谐, 属下这几个种的传统分类学结果是分类学家所公认的(杨光耀等, 1993)。因此利用这种标记技术产生的数据获得的结果较为合理, 说明这种标记可用于这几个竹种亲缘关系的研究。而属间的关系从聚类图上大致可以分为2个大类, 苦竹属(Pleioblastus)、矢竹属(Pseudosasa)与青篱竹属(Arundinaria)为1大类, 该类群又分为3个关系较近的分支。而巴山木竹属(Bashania)是单独的1类, 与这几种竹子的亲缘关系相对较远, 这一结果与尹佟明等和杨光耀等(个人交流)利用RAPD标记技术对巴山木竹的研究结果一致。从遗传距离看, 巴山木竹属作为1个单独的属是成立的。

|
图 2 POPGEN32软件生成的各竹种聚类结果图 Fig. 2 The dendrogram produced by POPGEN32 software forvarious bamboo species |
|
|
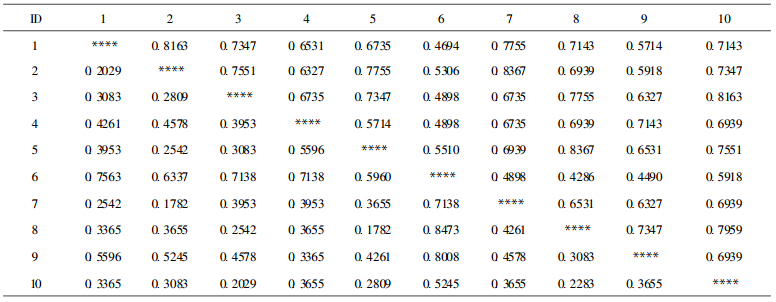
在传统的分类学研究中, 一般认为巴山木竹(Bashania fargesii)与苦竹属(Pleioblastus)亲缘关系较近, 有的学者则认为巴山木竹应归入苦竹属中, 本研究结果显示巴山木竹属(Bashania)与苦竹属(Pleioblastus)类群的关系比矢竹属(Pseudosasa)、青篱竹属(Arundinaria)有更近的遗传距离, 但相对于这两个类群而言, 巴山木竹属是1个有明显差别的类群。如果将聚类结果的第1和第2类群[包括矢竹属(Pseudosasd)、苦竹属(Pleioblastus) ]归并到青篱竹属中, 巴山木竹属能否归并及其在竹子分类系统中的位置需要进一步的研究和探讨。
在传统的竹子分类研究中, 茶秆竹(Arundinaria amabiliis)的去向一直是争论的热点, 中外竹类分类学者有主张将茶秆竹归入青篱竹属(Arundinaria)的, 有主张归入矢竹属(Pseudosasa)的, 陈守良等(1983)利用数量分类方法得到茶秆竹与矢竹关系较近, 得出茶秆竹应归入矢竹属的结论。王正平等(1980)也认为茶秆竹应并入矢竹属。本研究结果显示茶秆竹、福建茶秆竹是与矢竹属、苦竹属平行的1个类群, 而且矢竹与Arundinaria属的菲白竹、翠竹、苦竹属竹种的关系比其与茶秆竹的关系更近。由表 2看出, 矢竹与茶秆竹的Nei′s遗传距离为0.4261, 显示两者存在较大程度的分化。由于本研究中未采到青篱竹属的模式种, 因此对茶秆竹的归属还不能下定论。中国的茶秆竹与日本的矢竹存在较大的遗传差异, 本研究结果显示, 以苦竹属作为参照来看, 将其归入矢竹属是不妥当的。因此, 本研究不支持陈守良(1983)、王正平等(1980)的观点。
竹子特殊的开花习性使其分类远较有花植物其它类群困难, 属的分类尤其如此。竹子花果标本不易采到, 迫使研究者把目光转向营养体, 试图另辟蹊径以弥补这一不足。本世纪以来, 竹类植物营养体的多学科研究已经取得了大量成果, 部分已用于系统分类。但由于不同研究者对建立分类等级, 特别是属的标准取舍不一, 造成一些属的分合之争旷久不休(杨光耀等, 1993)。自然界中生物类群之间的形态差异是客观存在的, 它们不可能是完全同一的。系统分类的研究不仅仅依据某一形态特征的差异, 更重要的是所依据的形态差异应具有演化意义, 因此仅仅依据营养体性状进行研究具有很大的局限性。现代生物学的发展为这一问题的研究提供了更有效的方法, 其中根据在进化上有同源性的同源基因家族的进化关系推断各竹种间的系统关系是最有效的方法。仅根据本研究结果对青篱竹属分类上的疑问还不能给出确切答案, 因为种、属所跨越的遗传距离并没有确切的量的概念, 但本研究为从不同层面研究这几种竹子的亲缘关系提供了1个新的方向, 同时为这几个竹子系统学研究过程中合适序列片段的选择提供了借鉴。
与RAPD标记不同, 本研究使用了水稻的微卫星序列进行研究, 由于两者在进化上存在较近的关系, 所以这些引物在对竹子的研究中有一定的适用性。但我们无法确定相应的特异PCR产物, 只能根据扩增产物的重复性和微卫星片段的大小划定大致的范围, 因此还需通过测序进一步验证这些谱带的同源性。但本研究中关系明确种(如苦竹属)亲缘关系的验证, 说明这种方法还是有效的。本研究为竹子的研究指明了1个很好的方向, 利用水稻分子生物学的研究成果进行竹子相关领域的研究将是1个极富成果的研究方向。在植物系统发育研究过程中, 通常采用的基因位点包括叶绿体基因组的16S rRNA, 23S rRNA, psbA, psbD, psaB, psbC, psaA, rbcL, atpB, ndhA, atpA, ndhD, rpoB, rpoCI, rpoA, ndhF, rpoC2, matK基因等(汪小全等, 1997)。叶绿体基因组的内含子(Intron)区和基因间隔区(Intergenic spacers, IGS)比编码区存在更多的变异(Taberlet et al., 1991; Demesure et al., 1995; Perez de la Rosa et al., 1995), 这些DNA区段在植物的系统发育研究中应用也较为广泛。另外, 核基因组ITS区序列在植物的系统发育方面也有较广泛的应用。孙晔等(私人交流, 未发表)利用具有中等进化速率的ndh基因对竹子的系统发育进行研究, 结果发现不同竹种在该基因序列上存在较小的差异, 不能提供足够的系统发育信息。本研究的结果表明青篱竹属相关属除巴山木竹属外, 其余属亲缘关系相对较近, 因此在这些属的系统学研究中, 在序列的选择上应选择那些进化速率较快的基因序列。上述基因位点中, matK基因、叶绿体基因组的内含子(如trnintron)、叶绿体基因基因间隔区(如rp133-psaJ-trnP基因间隔区)及核基因组rDNA的ITS区有较快的进化速率, 所以在青篱竹属相关属的系统学研究中, 应从这些基因位点中进行选择。
本研究所采用的方法并不是严格意义上的SSR标记技术, 而是利用水稻的SSR引物在水稻基因组中进行随机扩增, 这一方法对比于RAPD标记在两个方面提高了扩增谱带的特异性, 1是引物序列较长, 2是与竹子基因组存在一定的同源性。根据近缘种的聚类结果可以看出, 这一方法适用于青篱竹属相关属亲缘关系的研究。但要获得更可靠的研究结果, 还需要对可能的微卫星扩增谱带进行测序验证。但开发微卫星引物的成本较高, 因此本研究也给出了一个在竹子中开发SSR的新途径。另外SSR标记是共显性的, 本研究将在竹子中获得的可能的微卫星序列作为显性标记进行处理, 由于不是将该标记作为群体遗传分析的手段, 如基因频率和等位基因数的估算等, 仅是用于作为聚类分析的数据来源, 所以这一处理对本研究的结果影响较小。综上所述, 本研究的结果是比较可靠的。
陈守良, 徐克学, 盛国英. 1983. 中国散生竹的数量分类和分类等级的探讨. 植物分类学报, 21(2): 113-119. |
耿伯介. 1982. 世界竹亚科各属的考订. 竹子研究汇刊, 1(1): 1-19. |
何平. 1998. 真核生物中的微卫星及应用. 遗传, 20(4): 42-47. |
王正平, 叶光汉. 1980. 关于我国散生竹的分类问题. 植物分类学报, 8(3): 283-290. |
汪小全, 洪德元. 1997. 植物分子系统学近五年的研究进展概况. 植物分类学报, 35(5): 465-480. |
温太辉. 1983. 论竹类起源. 竹子研究汇刊, 2(1): 1-8. |
杨光耀, 赵奇僧. 1993. 中国青篱竹属的整理. 竹子研究汇刊, 12(4): 1-7. |
邹喻萍. 1995. RAPD分子标记简介. 生物多样性, 3(2): 104-108. DOI:10.3321/j.issn:1005-0094.1995.02.010 |
Couch J A, et al. 1990. Isolation of DNA from plant high in polyphenolic. Plant Mol Biol Rep, 8: 8-12. DOI:10.1007/BF02668875 |
Demesure B, Sodzi N, Petit R J. 1995. A set of universal primers for amplification of polymorphic non-coding regions of mitochondrial and chloroplast DNA in plants. Mol Ecol, 4: 129-131. DOI:10.1111/j.1365-294X.1995.tb00201.x |
Gernano J, Klein A S. 1999. Species-specific nuclear and chloroplast single nucleotide polymorphisms to distinguish Picea glauca, P. mariana and P. rubens. Theor Appl Genet, 99: 37-49. DOI:10.1007/s001220051206 |
Grattapaglia D, Chaparro J X, Wilcox P et al. Mapping in woody plants with RAPD markers: Application to breeding in forestry and horticulture. In: proceedings of the symposium in aplication of RAPD technology to plant breeding, CSSA/ASHS/AGA, 1992, 37~40
|
Kimura M, Ohta T. 1973. Mutation and evolution at the molecular level. Genetics, 73(supplement): 19-35. |
Li J H, Bogle A L, Klein A S. 1997. Interspecific relationships and genetic divergence of the disjunct genus Liquidambar (Hamamelidaceae). Rhodora, 99: 229-240. |
Moore G, Devos K M, Wang Z, et al. 1995. Grasses, line up and from a circle. Current Biology, 5: 737-739. DOI:10.1016/S0960-9822(95)00148-5 |
Nei M, Roychoudhury A K. 1973. Sampling variances of heterozygosity and genetic distance. Genetics, 76: 379-390. |
Olmstead R G, Palmer J D. 1994. Chloroplast DNA systematics:a review of methods and data analysis. Am J Bot, 81: 1205-1224. DOI:10.1002/j.1537-2197.1994.tb15615.x |
Perez de la Rosa J, Harris S A, Farjon A. 1995. Noncoding chloroplast DNA variation in Mexican pines. Theor Appl Genet, 91: 1101-1106. |
Steele K P, Vilgalys R. 1994. Phylogenetic analysis of Polemoniaceae using nucleotide sequences of the plastid gene matk. Sys Bot, 19: 126-142. DOI:10.2307/2419717 |
Taberlet P, Gielly L, Pautou G, et al. 1991. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Mol Biol, 17: 1105-1109. DOI:10.1007/BF00037152 |