 2002, Vol. 38
2002, Vol. 38文章信息
- 张小全, 徐德应.
- Zhang Xiaoquan, Xu Deying.
- 温度对杉木中龄林针叶光合生理生态的影响
- EFFECTS OF TEMPERATURE ON THE PHOTOSYNTHETIC PHYSIOECOLOGY OF 18-YEAR-OLD CHINESE FIR(CUNNINGHAMIA LANCEOLATA(LAMB.)HOOK.)
- 林业科学, 2002, 38(3): 27-33.
- Scientia Silvae Sinicae, 2002, 38(3): 27-33.
-
文章历史
- 收稿日期:1999-04-21
-
作者相关文章


温度是影响植物光合生理生态最重要的环境因子之一。每种植物都须在一定温度范围内才能进行光合作用并有其最低、最适合和最高温度范围(Larcher, 1983)。国外从20世纪70年代就对树木光合生理生态对温度的响应进行了大量的研究和报道(Slayter et al., 1977a; 1977b; Berry et al., 1980; Kirschbaum et al., 1984; Battaglia et al., 1996), 并用于冠层光合和蒸散的模拟(Running et al., 1988; Kajfez-Bogataj, 1990; McMurtrie et al., 1989; 1990; Aber et al., 1992; Zhang et al., 1994; Sands, 1995)。近年来在国内对木本植物或树木也开展了少量的研究(贺东祥等, 1995; 王邦锡等, 1997; 胡新生等, 1997), 但多限于苗木。尽管许多学者都曾对杉木光合生理生态进行过研究测定(李竹林等, 1980; 齐金银, 1989; 郭继善, 1992; 吴立勋等, 1992; 陈洪, 1995; 范少辉等, 1996; 张小全等, 1999), 但未就温度对其光合生理生态的影响进行过系统的研究。本文系统研究了杉木中龄林针叶净光合、蒸腾、气孔导度和暗呼吸等对温度的响应。
1 样地基本情况研究试验样地设于江西省分宜市亚热带林业实验中心年珠实验林场, 地理位置114°33′47″E, 27°34′41″N, 属罗霄山脉北端武功山支脉, 是杉木分布的中带中部产区。地带性植被为常绿阔叶混交林。气候属亚热带湿润气候区, 四季分明, 气候温和, 日照充足, 雨量充沛。年均气温16.8℃, 极端最高温39.9℃, 极端最低温-8.3℃, 年积温5355℃; 年降水量1590 mm, 年蒸发量1504 mm; 年均日照1657 h; 太阳辐射年总量约487 kJ·cm-2; 年均相对湿度80%;无霜期270 d。试验样地为杉木人工纯林, 林龄18 a, 地位指数16, 株行距2.0 m×1.5 m, 平均胸径13.8 cm, 平均树高12.6 m, 平均冠幅2.89 m, 枝下高7.52 m。海拔240~270 m, 坡向南偏东20°, 坡度20°。成土母质为砂页岩, 土壤属黄棕壤, 土层厚度70~100 cm。
2 野外实验方法在样地内根据林分平均胸径、平均树高、平均冠幅和冠长, 选取1株标准木, 分别从树冠中部向南伸展的侧枝上选择当年生、1 a生和2 a生(包括2 a生以上, 以下简称2 a生)代表性样枝, 分别于1998年生长盛期(5月下旬)、中期(7月下旬)和后期(10月中旬), 在大气CO2浓度(350±10) μL·L-1左右、相对湿度70%±5%左右、光合有效辐射通量密度(PPFD)为400、800、1200 μmol·m-2s-1左右条件下测定不同叶温下针叶的净光合速率(Pn)、蒸腾速率(E)和气孔导度(gs)。所用仪器为LiCor-6200光合测定系统, 但对其4 L叶室进行了改装, 在底部加装了散热板并通过一温控器来控制和调节叶室内气温。测定温度范围10℃~40℃ (夏季15℃~40℃)。在每一温度下至少稳定5 min以使叶片充分适应新的温度条件, 当光合速率稳定后, 再进行测定(Battaglia et al., 1996)。测定是在太阳辐射较稳定的天气(无云晴天或阴天)条件下进行, 用不同厚度白色纱布覆盖叶室, 并辅以镝灯来达到和稳定辐射强度。辐射感应头被移至叶室内部中央并与针叶处于同一高度, 以保证测定的辐射强度与针叶接收的实际强度一致。根据我们测定, 针叶光饱和点在700 (2 a生) ~1800 (当年生) μmol·m-2s-1, 由于在高强辐射下, 叶温增加较快而难以稳定, 故试验在较低的辐射强度下进行。当地实际大气湿度一般均在70%以上, 调节通过干燥剂的气体流量, 以使叶室内相对湿度维持在70%±5%范围。通过人工用细管吹气的方法来增加叶室内的CO2浓度并利用系统内的CO2吸收器(Scrubber)来快速改变叶室内CO2的浓度, 使每次测定时叶室内CO2浓度控制在(350±10) μL·L-1。测定从早晨(7点左右)开始温度由低到高。早晨较低的气温可使叶室内的气温更容易降低到较低水平; 而早晨较高的大气湿度在降温后也可达到70%左右。必要时, 在整个测定过程中可打开叶室, 使其重新获得外界的温、湿度环境, 然后重新调节控制所需的叶室环境。
用相似的方法控制叶室环境, 用遮光法测定针叶暗呼吸对温度的响应。
由于对叶室环境的控制十分困难, 因此仅在每个季节于每一叶龄级选择一代表性样枝进行测定。
在1998-05~1998-10全部测定期间雨量充沛, 未发生土壤水分亏缺, 因此可以认为土壤水分供应充足。上述测定是在叶室中以单位活体小枝为单位进行的。杉木针叶为披针形片状体, 且单面有气孔, 故小枝针叶面积可采用复印称重法测定, 即将测定枝上的针叶全部剪下, 用透明胶带粘贴于复印纸上复印, 将复印出的针叶剪下称重, 根据单位复印纸的重量计算测定枝上的叶面积。
3 研究结果 3.1 杉木针叶净光合速率(Pn)对温度的响应光合作用和其它生化过程一样, 其速率显著地受温度的影响(沈允纲等, 1992)。同时温度的变化还会引起空气饱和水汽压差的变化, 从而影响气孔张开度, 对叶片与大气之间的CO2和水汽交换产生间接影响。一些研究指出, 光合速率随叶片温度的变化呈抛物线型关系, 可用下述经验方程描述二者之间的关系(Sall et al., 1994; Battaglia et al., 1996) :
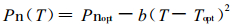
|
(1) |
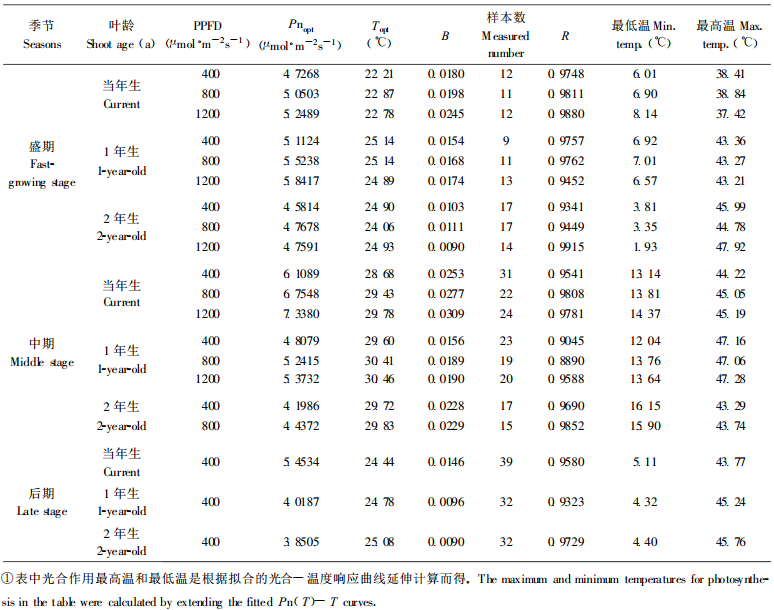
式中, Pn (T)是温度为T时的Pn; b为参数, 代表抛物线的曲率, 即对温度变化的敏感度; Topt表示光合作用的最适温度, Pnopt表示Topt时的Pn。将杉木生长盛期、中期和后期树冠中部测定的不同年龄针叶在不同温度下的Pn, 用(1)式进行拟合, 取得较好效果。拟合结果见表 1和图 1。为便于不同季节的比较且由于篇幅所限, 图 1仅显示了400 μmol·m-2s-1左右PPFD下的测定结果。
|
|

|
图 1 不同季节不同年龄杉木中龄林针叶净光合速率(Pn)对温度的响应(正常大气CO2浓度, 400 μmol·m-2s-1 PPFD和70%±5%相对湿度) Fig. 1 Temperature responses of net photosynthetic rate (Pn) for different age of Chinese fir shoots in 3 growing seasons with normal ambient CO2, 70%±5% relative humidity and 400 μmol·m-2s-1 PPFD  当年生Current shoot; 当年生Current shoot;  1 a生1-year-old shoot; 1 a生1-year-old shoot;  2 a生2-year-old shoot. 2 a生2-year-old shoot.
|
图 1表明, 杉木中龄林针叶Pn对温度变化十分敏感。随着温度的升高, Pn增加, 到达一定温度后, 虽然总光合速率仍维持在较高水平, 但可能由于光呼吸大幅度上升, Pn急剧下降。有研究表明, 光呼吸与暗呼吸一样随温度的增加而增强(高煜珠等, 1982)。
表 1表明, 不同生长季节最适温度范围不同, 反映了针叶对环境温度的适应特征。在生长盛期和后期针叶Pn最适温度为25℃左右, 但生长盛期的当年生叶例外, 只有22~23℃。生长中期由于环境温度上升, 针叶Pn的最适温度也上升了5℃左右, 达29~31℃。光合作用的温度范围也存在季节差异, 在生长盛期和后期, Pn的最低温度为5℃左右, 而在生长中期为15℃左右, 但最高温度均为45℃左右(盛期当年生叶除外)。虽然这个温度范围是通过模拟曲线的延伸而来, 可能存在误差, 但仍可看出一定的规律性。从经验方程中参数b的值看, 生长中期最大, 说明中期针叶光合作用对温度变化的反应最剧烈。
就不同年龄针叶对温度的响应而言, 当年生新针叶与老叶存在一定差异, 特别是在生长盛期。不但新生叶Pn的最适温度明显低于老叶2~3℃, 而且其最高温度仅38℃左右, 比老叶低5℃以上, 温度范围明显比老叶窄。随着当年生叶的生长、组织的成熟和对环境适应能力的增强, 到生长中、后期, 当年生针叶与1 a生、2 a生针叶光合作用的温度范围和最适温度已无明显差异(表 1)。Battaglia等(1996)对4 a生桉树(Eucalyptus globulus)的研究也表明, 正在生长的叶、当年生成熟叶片和老叶片的光合最适温度有显著差异。这种差异可能与生长呼吸而不是与光合机构的适应能力有关, 因为随着温度的上升, 呼吸速率的增加速度一般比羧化速率要大(Berry et al., 1980)。通常, 幼嫩组织呼吸速率大于老组织, 正在生长的组织的呼吸速率大于成熟组织。5、6月份是杉木新生针叶快速生长期, 不但具有较高的生长呼吸速率, 而且呼吸速率随温度而剧烈变化, 导致上述盛期当年生针叶Pn对温度的响应特征。
上述光合速率对温度的响应实际上是对温度变化的瞬时响应(数分钟)。当环境温度在短时间内偏离最适温度时, 光合活性降低, 但当恢复到最适温度后, 光合速率又会恢复到最大值。然而当长时间偏离最适温度, 光合作用和呼吸作用都会对温度的变化产生适应(Read et al., 1990; Battaglia et al., 1996), 使最适温度发生变化。高环境温度可使光合最适温度提高, 低环境温度使光合最适温度降低(高煜珠等, 1982)。因此这种瞬时响应也是基于对环境温度的适应。有人认为, 光合作用的最适温度接近于叶片前10 d所处环境中最高温度的平均值(Slayter et al., 1977a; 沈允纲等, 1992)。Koner和Diemer (1984)认为植物光合最适温度接近晴天平均叶温。Battaglia et al. (1996)研究认为, 当年生成熟叶光合最适温度与测定前一个星期内的平均温度线性相关。从我们的测定结果与当地气象资料对比看, Pn的最适温度更接近于测定前5 d日最高气温的平均值。
3.2 杉木中龄林针叶气孔导度(gs)对温度的响应气孔是植物与大气进行水汽和CO2交换的通道, 气孔的开闭会对植物叶片的光合和蒸腾作用产生影响, 气孔导度则是反映气孔行为最重要的生理指标。然而气孔的开放与关闭是多种因素共同作用的结果, 如胞间CO2浓度(Ci)、空气相对湿度(Rh)、温度等。以生长后期为例(图 2-A~C), 杉木针叶gs对温度的响应呈抛物线型, 但其取得最大值的温度比光合最适温度要低得多。由于gs是叶面相对湿度或叶面-空气水汽压饱和差(VPD)、胞间CO2浓度(Ci)和光合速率的函数(Ball et al., 1987; Leuning et al., 1995), 温度对gs的影响可能是通过VPD变化而引起的间接影响, 同时受光合作用引起的Ci变化的作用。随着温度的增加, VPD增大(图 2-D~F), 由于气孔响应失水而收缩, 失水速率与VPD成正比(Aphalo et al., 1991; Sheriff, 1984), 因而会限制气孔的开放。但另一方面, 随着温度的增加, Pn迅速增大, Ci降低, 这又会促进气孔的开放。由于在低温阶段, VPD较小, 对气孔的限制作用较小, Pn的增加对气孔开放的促进作用占主导地位, 因此gs迅速增加。随着温度的继续增加, VPD逐渐增大, 其引起的限制作用也增强, 当这种限制作用与光合作用引起的促进作用相平衡时, gs达到最大值。以后, 尽管光合速率仍在增加, 但VPD的作用占据主导地位, gs呈迅速下降趋势(图 2-A~F)。

|
图 2 生长后期杉木针叶蒸腾速率(E)、气孔导度(gs)对温度的响应(正常大气CO2浓度、相对湿度70%±5%、PPFD 400 μmol·m-2s-1)及饱和水汽压差(VPD)与温度的关系 Fig. 2 Responses of transpiration rate (E) and stomatal conductance (gs) to temperature for different age of Chinese fir shoots and VPD in relation to temperature, with normal ambient CO2 content, relative humidity 70% and PPFD 400 μmol·m-2s-1 ▲E, □gs, ●VPD (水汽压差Vapour saturated deficient). |
叶片蒸腾作用受温度、VPD、叶界面层阻力、气孔阻力及土壤水分供应等内外部因子的影响(Kozlowski et al., 1991)。在本试验条件下(充足的土壤水分供应, 正常大气CO2浓度(350±10) μL·L-1、相对湿度70%±5%、PPFD 400 μmol·m-2s-1左右), 针叶E对温度变化的响应与gs和Pn相似, 但其达最大E的温度比gs高, 与光合最适温度相近(图 1-A~C和图 2-A~C)。虽然气孔是水汽从叶内向外扩散的主要通道, 气孔阻力则是水汽从叶内向外扩散的唯一重要限制因素。但是由于蒸腾作用还受VPD的影响, 随VPD的增大而增加(Kozlowski et al., 1991), 使E对温度的响应复杂化。当温度从较低温度开始上升时, gs增加, VPD增大, 故E增加。当gs达到最大值后开始剧烈下降时, 由于VPD继续增加, 促进叶片蒸腾; 加之角质层蒸腾速率不受gs影响而随温度增加而直线上升(Sheriff, 1984), 因而E仍继续增加。随着温度的继续增加, VPD增大使气孔部分或全部关闭, E开始下降(图 2-A~F)。
3.4 杉木针叶暗呼吸对温度的响应树木叶片的呼吸作用受多因子的影响, 包括叶片所处的光环境、温度、叶龄等(Hagihara et al., 1991; Kramer, 1985)。对生长盛期杉木树冠中部不同年龄针叶暗呼吸的测定表明(图 3-A~C), 暗呼吸速率随温度的增加呈指数上升趋势, 用下述指数方程可较好地表达这种关系(Landsberg, 1986) :
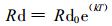
|
(2) |

|
图 3 生长盛期杉木中龄林针叶暗呼吸对温度的响应 Fig. 3 Temperature dependence of dark respiration of Chinese fir shoot in fast-growing stage |
式中, Rd为暗呼吸速率; Rd0为T=0℃时的呼吸速率; T为叶温; k为参数。用(2)式进行拟合得:
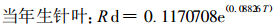
|
(3) |

|
(4) |
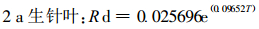
|
(5) |
图 3-D和式(3) ~ (5)表明, 不同针叶年龄, 暗呼吸有明显的差别。当年生针叶, 其暗呼吸速率明显高于1 a生和2 a生老叶。这是因为5月下旬是生长高峰, 幼嫩组织呼吸速率大于老组织, 生长组织的呼吸速率大于成熟组织。此外, 不同年龄针叶暗呼吸速率的差异还可能与其所处的光环境有关, 从冠层上部到下部叶片的暗呼吸速率随叶片接受辐射的减少而降低(Hagihara et al., 1991)。老叶尤其是2 a生及2 a生以上针叶一般处于树冠的里层和下层, 受光少, 当年生叶则处于树冠顶层或外围, 光环境优越。
通常用温度增加10℃化学反应速度增加的倍数(Q10)来表示生物过程对温度变化的反应。根据上述方程计算, 在生长盛期, 杉木中龄林当年生、1 a生和2 a生针叶暗呼吸的Q10分别为2.43、2.68和2.62。大多数研究测定的Q10略高于2 (Landsberg, 1986), 然而Q10的值因环境温度而异。Hagihara等(1989)的测定表明, 暗呼吸的Q10在1.4~3.4之间, 冬季较高而夏季较低, 表现出明显的季节变化特征; Q10与月平均气温呈线性相关。Larcher (1983)综合了许多研究结果后认为, 植物呼吸作用的Q10在5~25℃时为2.0, 低于5℃时为3.0或更高, 在25~30℃时降到1.5以下。Lyons (1973)认为, 植物呼吸作用Q10的这种变化与酶的活化能的转变有关(Hagihara et al., 1991)。遗憾的是, 本试验未对叶片暗呼吸Q10的季节变化进行测定。
尽管呼吸作用也有其最适温度范围(苘辉民, 1994), 但我们没有测定到呼吸速率开始下降的温度, 可能与测定的时间较短有关。超过最适温度, 在短时间内呼吸速率仍可继续增高, 但时间延长后, 呼吸速率就会急剧下降(苘辉民, 1994)。
4 结论在其它环境因素相对稳定的条件下, 本试验研究表明:
针叶净光合速率(Pn)对温度的响应呈抛物线型, 不同季节Pn的最适温度不同, 在生长盛期(5月下旬)和后期(10月中旬)为25℃左右, 在生长中期30℃左右, 但生长盛期当年生针叶例外, 其Pn最适温度仅22℃~23℃。Pn的最适温度与测定前5 d平均最高气温接近。
针叶气孔导度(gs)和蒸腾速率(E)对温度的响应与Pn相似, 但gs达最大值的温度比Pn低。温度对gs的影响可能源于叶面-大气水汽压差(VPD)的变化和光合作用引起的胞间CO2浓度的改变。温度对E的影响与gs和VPD的变化密不可分。
针叶暗呼吸速率(Rd)随温度增加呈指数增加, 但由于生理活性和所处光环境的差异, 当年生针叶Rd明显高于1 a生和2 a生老叶。
杉木针叶光合生理生态对温度变化的长期适应有待进一步研究。
陈洪. 1995. 杉木幼林若干光合特性的研究. 福建林业科技, 22(3): 69-72. |
范少辉, 沈国舫, 程仲辉, 等. 1996. 不同立地条件不同林龄杉木人工林生理特性的研究Ⅱ.光合特性. 林业科学研究, 9(mem): 60-65. |
高煜珠, 王忠. 1982. 关于光呼吸与光合作用关系的研究Ⅱ.环境因素对光合作用的影响及其与光呼吸的关系. 植物生理学报, 8(4): 373-382. |
郭继善. 1992. 利用14CO2研究杉木的光合特性. 南京林业大学学报, 16(4): 86-89. |
贺东祥, 沈允钢. 1995. 几种常绿植物光合特性的季节变化. 植物生理学报, 21(1): 1-7. DOI:10.3321/j.issn:1671-3877.1995.01.011 |
胡新生, 刘建伟, 王世绩. 1997. 四个杨树无性系在不同温度和相对湿度条件下净光合速率的比较研究. 林业科学, 33(2): 107-116. DOI:10.3321/j.issn:1001-7488.1997.02.002 |
齐金银. 1989. 杉木北缘产地主要气候生态因子与杉木生理活动和径向生长关系初探. 植物学报, 31(6): 468-476. |
沈允钢, 许大全. 1992. 植物生理与分子生物学Ⅹ Ⅴ Ⅲ光合机构对环境的响应与适应. 北京: 科学出版社, 225-235.
|
苘辉民. 1994. 植物生理学. 北京: 北京农业大学出版社.
|
王邦锡, 王辉, 黄久常. 1997. 沙拐枣同化枝的光合作用和呼吸作用对生长季节、光照强度、高温和干旱的响应. 林业科学, 33(1): 19-24. DOI:10.3321/j.issn:1001-1498.1997.01.004 |
吴立勋, 徐世凤. 1992. 杉木、檫树光合特性研究. 湖南林业科技, (2): 5-10. |
张小全, 徐德应, 赵茂胜, 等. 2000. CO2增长对杉木中龄林针叶光合生理生态的影响. 生态学报, 20(3): 390-396. |
Aber A D, Federer C A. 1992. A generalized, lumped-parameter model of photosynthesis, evapotranspiration and net primary production in temperate and boreal forest ecosystems. Oecologia, 92: 463-474. DOI:10.1007/BF00317837 |
Aphalo P J, Jarvis P G. 1991. Do stomata respond to relative humidity?. Plant, Cell & Environ, 14: 127-132. |
Ball J T, Woodrow L E, Berry J A.A model predicting stomatal conductance and its contribution to the control of photosynthesis under different environmental conditions.In: Binggins L J(ed).Progress in photosynthesis research Vol.Ⅳ.Netherlands: Martinus Nijhoff, Dordrecht, 1987, 221~224
|
Battaglia M, Beadle C, Loughhead S. 1996. Photosynthetic temperature responses of Eucalyptus globulus and Eucalyptus nitens. Tree Physiology, 16: 81-89. DOI:10.1093/treephys/16.1-2.81 |
Berry J, Bjorkman O. 1980. Photosynthetic response and adaptation to temperature in higher plants. Annu Rev Plant Physiol, 31: 491-543. DOI:10.1146/annurev.pp.31.060180.002423 |
Hagihara A, Hozumi K.Respiration.In: Raghavendra A S(ed).Physiology of Trees.New York: John Wiley & Sons, INC, 1991, 87~110
|
Hagihara A, Paembonan S A, Hozumi K. 1989. Respiration measurement of a hinoki forest-tree over a three-year-period. Trans Mtg Jpn For Soc, 100: 443. |
Kajfez-Bogataj L. 1990. Photosynthetic model for predicting net willow stand production. Agric For Meteorol, 50: 55-85. DOI:10.1016/0168-1923(90)90138-V |
Kirschbaum M U F, Farquhar G D. 1984. Temperature dependence of whole-leaf photosynthesis in Eucalyptus pauciflora Sieb.ex Spreng. Aust J Plant Physiol, 11: 519-538. |
Koner C H, Diemer M. 1984. CO2 exchange in the alpine sedge Carex curvula as influenced by canopy structure, light and temperature. Oecologia, 53: 98. |
Kozlowski T T, Kramer P J, Pallardy S G. 1991. The Physiological Ecology of Woody Plants. San Diego: Academic Press.
|
Kramer P T著, 汪振儒译.木本植物生理学.北京: 中国林业出版社, 1991
|
Larcher W. 1983. Physiological Plant Ecology(corrected printing). Berlin: Springer-Verlag.
|
Landsberg J J. 1986. Physiology Ecology of Forest Production. London: Academic Press.
|
Leuning R. 1995. A critical appraisal of a combined stomatal-photosynthesis model for C3 plants. Plant Cell & Environ, 18: 339-355. |
McMurtrie R E, Landsberg J J, Linder S.Research priorities in field experiments on fast-growing tree plantations: implications of a mathematical model.In: Pereira J S, Landsberg J J(eds).Biomass Production by fast-Growing Trees.Kluwer, Dordrecht, 1989, 181~207
|
McMurtrie R E, Rook D A, Kelliher F M. 1990. Modelling the Yield of Pinus radiata on a site limited by water and nitrogen. For Ecol M anage, 30: 381-413. DOI:10.1016/0378-1127(90)90150-A |
Read J, Busby J R. 1990. Comparative responses to temperature of the major canopy species of Tasmanian cool temperature rainforest and their ecological significance.Ⅱ.Net photosynthesis and climate analysis. Aust J Bot, 38: 185-205. DOI:10.1071/BT9900185 |
Running S T, Coughlan J C. 1988. A general model of forest ecosystem processes for regional applications.Ⅰ.Hydrologic balance, canopy gas exchange and primary production processes. Ecol Model, 42: 125-154. DOI:10.1016/0304-3800(88)90112-3 |
Sall T, Pettersson P. 1994. A model of photosynthetic acclimation as a special case of reaction norms. Theor Biol, 166: 1-8. DOI:10.1006/jtbi.1994.1001 |
Sands P J. 1995. Modelling canopy production.Ⅱ.From single-leaf photosynthetic parameters to daily canopy photosynthesis. Aust J Plant Physiol, 22: 603-614. |
Sheriff D W. 1984. Epidermal transpiration and stomatal responses to humidity :Some hypotheses explored. Plant, Cell & Environ, 7: 669-677. |
Slayter R O, Ferrar P J. 1977a. Altitudinal variation in the photosynthetic characteristics of snow gum, Eucalyptus pauciflora Sieb.Ex Spreng.Ⅱ. Effect of growth temperature under controlled conditions. Aust J Plant Physiol, 4: 289. |
Slayter R O, Ferrar P J. 1977b. Altitudinal variation in the photosynthetic characteristics of snow gum, Eucalyptus pauciflora Sieb.Ex Spreng.Ⅴ.Rate of acclimation to an altered growth environment. Aust J Plant Physiol, 4: 595. |
Zhang Y. 1994. A process-based growth model for young red pine. Forest Ecology and Management, 69: 21-40. DOI:10.1016/0378-1127(94)90217-8 |