 2002, Vol. 38
2002, Vol. 38文章信息
- 吴小芹, 何宗明, 魏玉莲.
- Wu Xiaoqin, He Zhongming, Wei yulian.
- 我国松杉球壳孢生长适应性分化研究
- STUDY ON GROWTH ADAPTABILITY DIFFERENTIATION OF SPHAEROPSIS SAPINEA IN CHINA
- 林业科学, 2002, 38(2): 78-82.
- Scientia Silvae Sinicae, 2002, 38(2): 78-82.
-
文章历史
- 收稿日期:2001-02-16
-
作者相关文章


2. 福建农林大学林学院 南平 353001
2. Forestry College, Fujian Agriculture and forestry university Nanping 353001
松杉球壳孢[ Sphaeropsis sapinea(Fr. : Fr.) Dyko &Sutton]普遍分布于世界各地, 主要引起针叶树枯梢、枝干溃疡流脂、根颈腐烂和木材蓝变等多种症状。我国自20世纪70年代以来, 相继有10多个省份报道了该病菌的发生与为害(吴小芹, 1999)。从目前情况看, 松杉球壳孢在我国发生地域广, 为害寄主多, 各地发病程度不一。为了解该病菌在与寄主和环境的相互作用过程中, 是否可能产生一定的适生性变异以适应生境的变化, 作者就来自我国不同地区针叶树上的松杉球壳孢对温度、培养基质及其基质酸碱度的适应效应进行了研究, 并运用高次多项式数学模型拟合方法对各菌株的适应性进行了数量化的分析比较, 以期为了解该病菌的群体分化状况和进一步深入揭示松树枯梢病在我国的分布特点及流行规律提供参考依据。
1 材料和方法 1.1 菌株来源供试的49 ~ 55个菌株来自我国13个省份, 北起黑龙江哈尔滨, 南至广东雷州半岛; 寄主树种包括16种松树和其它2种针叶树; 病样症状主要为枯梢, 其次为树干流脂溃疡; 其具体地理来源和采样寄主见参考文献(吴小芹, 2000)。
1.2 病菌生长适应性试验 1.2.1 对营养基质的生长适应性将菌龄一致的各菌株菌落圆片(直径5 mm), 分殖于PDA、Czapek、红松(Pinus koraiensis)松针煎汁培养基(HA)、马尾松(P.massoniana)松针煎汁培养基(MA)、湿地松(P. elliottii)松针煎汁培养基(SA)和玉米粉培养基(CMA)上并置25 ℃下无光照培养。每一处理设置3个重复。接菌24 h、48 h、72 h、96 h和120 h后分别测定各菌落生长状况。
1.2.2 对温度的生长适应性将各菌株菌落圆片(直径5 mm)移殖在PDA平板上, 分置于10 ℃、15 ℃、20 ℃、25 ℃、30 ℃和35 ℃等6种系列温度下无光照培养。生长速率测定方法与时间同上。
1.2.3 对pH值的生长适应性用HCl和NaOH在pHS -25型酸度计(上海雷磁仪器厂)上将供试PDA培养基的pH值分别调成pH3、4、5、6、7和8等6种酸度, 将各菌株菌龄一致的菌落圆片(直径5 mm)分别移殖至6种pH值的PDA上, 置25 ℃下无光照培养。生长速率测定方法同上。
1.3 数据分析将松杉球壳孢在系列温度和系列pH值下培养0 ~ 72 h的生长量, 采用高次多项式回归数学模型拟合方法, 构建出每一地理来源菌株的最适生长温度和最适生长pH值。
1.3.1 高次多项式数学模型拟合方法用郎奎健、唐守正编制的程序(BDXSHG. BAS) (郎奎健等, 1989)分别拟合各菌株的生长量与温度和pH值关系的高次多项式回归数学模型。经相关系数比较, 分别采用4次和3次多项式回归数学模型可较好拟合各菌株0 ~ 72 h的生长量随温度以及pH值变化的曲线。两模型分别为: 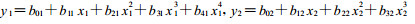
式中: y1、y2为某菌株生长量; x1为温度, x2为PH值; b01 ~ b41和b02 ~ b42为模型参数(根据所输入的某一菌株在不同温度或pH值下72 h的生长量由程序求出)。
1.3.2 最适生长温度或pH值的计算将上述模型求一阶导数, 分别如下:
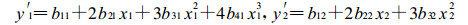
|
令 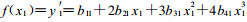
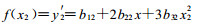
当f(x)=0时, y有一个极大值, 即最适温度或最适pH值时的生长量。用Newton法(钱焕延, 1988)求f(x)=0时的x值(初值选择在真菌的适生范围内)。此x值即为最适生长温度或最适生长pH值。
2 结果与分析 2.1 病菌群体在不同培养基质上营养生长适应性比较S. sapinea供试菌株在6种培养基质上72 h的生长状况表明, PDA是适于其生长的理想基质。在PDA上, 各菌株的营养生长差异不大; 除F1(福建湿地松)和J4(江苏黑松)生长较慢外, 其余菌株均生长较快(直径70 ~ 90 mm)。在Czapek上, 各菌株的生长速度和菌落形态变化较大; 其中GD2生长最快(直径90 mm), A1、D1、F6和F7生长受抑(< 20 mm), 说明供试菌株在利用矿质营养和微量元素(K、P、S、Mg、Fe)方面存在显著的适应性差异。在松针煎汁培养基中, 除J10(江苏火炬松)在SA上生长较慢外(直径17 mm), 其它各菌株在SA和MA上的生长整体趋缓, 波动不显(直径30 ~ 44 mm); 在HA上各菌株生长呈现一定差异, A1、D1、F7、J5、J3和S4等生长较快(直径55 ~ 69 mm), 而其它菌株次之。在CMA上, 菌落稀薄, 除J9、JX1、JX2和ZJ1生长扩展较慢外(直径45 ~ 54 mm), 多数菌株直径达55 ~ 69 mm。
2.2 病菌群体在系列温度下营养生长适应性比较从总体上看, 松杉球壳孢在10 ℃时即可生长, 并随着温度的递增, 生长量渐进上升, 直至25 ~ 30 ℃达到最大值; 从30 ℃至35 ℃, 生长量陡然下降(图 1)。多数菌株对系列温度的生长适宜顺序为: 25 ℃> 30 ℃>20 ℃>15 ℃。而从各个菌株上看, 不同菌株对系列温度的生长适应性存在着明显差异(见图 1和表 1)。在35 ℃时, 多数菌株均生长不好, 平均生长量仅为11.9 mm, 但来自广东马尾松上的GD2却仍可较好地生长, 其生长量达到55 mm, 超出平均水平的5倍; 而来自江苏红松和日本冷杉上的J20和J23以及来自云南高原加勒比松、海岸松和墨西哥变种白松上的YN4、YN5和YN6等在此温度下几乎不能生长。在30 ℃下, A1、F2、F5、F8、GD2、H1、J3、J5 -6、J13和JX2等菌株的生长量可达80 ~ 90 mm, 而其它菌株的生长量多介于50 ~ 80 mm之间, YN4和来自浙江乔松上的ZJ2生长量仅36 ~ 37 mm。在25 ℃、20 ℃和15 ℃下, 各菌株生长量的波动范围分别为61 ~ 90 mm, 43 ~ 76 mm和16 ~ 41 mm, 平均变幅极差均达25 ~ 30 mm左右。10 ℃下, 供试菌株的平均生长量为10.9 mm, 但J23、S4 (江苏日本赤松)、YN1和YN6的生长量略大些达14 ~ 15 mm, 而GD2和来自江苏连云港海岸松、湿地松和黑松上的J2、J3、J4以及YN4等却在10 ℃下几乎不长。研究表明, 与其它菌株比较, GD2的营养生长明显耐高温(35 ℃)而不耐低温(10 ℃); J20、J23、YN5和YN6等不耐高温; J2、J3、J4等不耐低温; 而YN4既不耐高温也不耐低温。

|
图 1 S. sapinea各菌株在系列温度下的总生长趋势(PDA, 72 h) Fig. 1 Growth of S. sapinea at different temperature on PDA (72 h) |
|
|

经用4次多项式回归数学模型拟合供试菌株0 ~ 72 h的生长量随温度变化的曲线, 构建出各菌株的最适生长温度, 见表 1。49个菌株的平均最适生长温度为26.8 ℃, 其中最低值为23.1 ℃, 最高值为29.6 ℃, 相互间最大差值达6.5 ℃, 说明各菌株的最适生长温度存在明显差异。
从地理来源来看, 来自云南、黑龙江、河南和陕西的菌株其生长最适温较低, 来自福建、江西、江苏、湖北和安徽等的菌株生长最适温较高; 但各菌株生长最适温的地理区划特征并不十分绝对, 如来自浙江乔松上的ZJ2菌株其生长最适温在供试菌株中是最低的(23.1 ℃)。从寄主树种来看, 来自湿地松和部分火炬松的菌株其最适生长温度较高些, 但树种主导作用似不明显。将地理区域与寄主树种对供试菌株的影响进行综合分析, 则地理区域的影响似乎大于寄主树种。如寄主同为火炬松, 来自河南和陕西的菌株(HN1和SX1)其生长最适温度相对低(25 ℃左右)于来自福建的菌株(F5和F7, 29 ℃左右)。
2.4 病菌群体在系列pH值下营养生长适应性比较从总体生长情况看, 供试55个菌株对系列pH值的适应范围较宽(图 2)。各菌株在pH3 ~ 8下均可生长, 其中pH5 ~ 6是大部分菌株生长较为适宜的范围。但从对每一pH值的生长反应来看, 供试菌株表现出一定差别。

|
图 2 S. sapinea各菌株在系列pH值下总生长趋势(PDA, 72 h) Fig. 2 Growth of S. sapinea at different pH on PDA (72 h) |
由图 2可见, 在总生长趋势曲线中, 除pH5外, 每个pH值的散点区域均较宽。在pH3和pH8时, 这些差别呈现得较为明显(图 2和表 2)。pH3时, 来自江苏红松的J20生长较慢(直径30 ~ 40 mm); 多数菌株生长居中(直径41 ~ 70 mm); D2、F6、F7、J8、YN1、YN4、YN5和YN6生长较快(直径71 ~ 80 mm); 而来自广东的GD2和LZ1则生长迅速(直径>85 mm)。由此可见, J20不耐酸, 而后2组菌株明显较耐酸。当pH8时, LZ1仍生长最快(直径>85 mm), 说明该菌株既耐酸又适碱; 而GD2与多数菌株的生长量接近, 不具明显优势(直径70 mm左右); F2、J2、J15和J19等则生长较慢(直径 < 60 mm)。
|
|
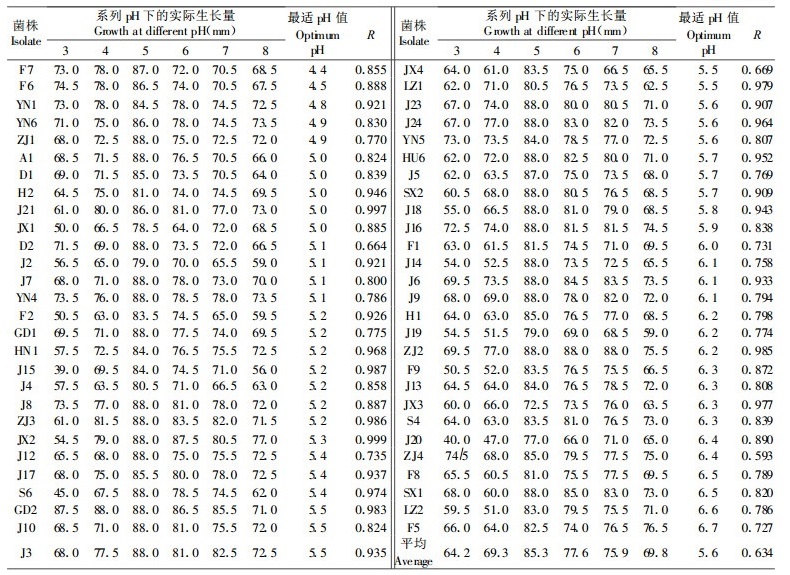
经用3次多项式回归数学模型拟合供试55个菌株0 ~ 72h的生长量随pH值变化的曲线, 构建出各菌株营养生长的最适pH值(表 2)。55个菌株的平均最适pH值为5.6, 其中最低值为pH 4.4(F7), 最高值为pH6.7(F5)。这表明各菌株的最适生长pH值存在一定差异。F6、F7、YN1、YN6、A1、和D1等的最适pH值较低(pH 4.4 ~ 5), 这几个菌株在培养性状上较相似(吴小芹, 2000), RAPD分析的分子树状图将它们聚在同一分枝内(吴小芹, 2000), 其中F6、F7、A1和D1还在同一个营养体亲和群(吴小芹, 2000)。而F5、F8、LZ2和SX1等的最适pH值较高(pH>6.5)。各菌株的最适生长pH值与地理来源和寄主树种似乎无明显关系。
3 结论和讨论来自我国不同地区不同寄主上的松杉球壳孢对不同或同一培养条件的生长反应存在较大变化。
在六种培养基中, 除MA和SA外, 各菌株在其余四种培养基上的生长均表现出明显的差异, 尤其在Czapek上, 说明菌株间在对营养的适应性上存在着明显的分化。
供试各菌株对温度的适应性变化表明, 虽然多数菌株的适温范围较宽, 但它们在对极端温度的反应和对最适温度的要求上还是显示出了较大的差别。来自云南高原的加勒比松和海岸松或来自江苏红松和日本冷杉上的菌株不耐高温, 而来自华南及其东南沿海地区的某些菌株在低温下生长不好。各菌株生长最适温度的变化似乎与地理来源有一定关系, 但也并不排除寄主及其小生境的影响。如来自同一地理区域(浙江杭州)不同寄主(晚松、乔松、日本五针松和马尾松)上的4个菌株其生长最适温也有差异(23.1 ~ 26.9 ℃)。将各菌株的最适生长温度与它们对几种松树实生苗的致病力结果(吴小芹, 2000)进行比较分析发现, 一些最适生长温度较高(>28 ℃)的菌株其致病力也往往较强。
松杉球壳孢对基质pH值的生长效应也存在着菌株间的差异, 尽管这些差异与地理来源或寄主树种以及致病力大小之间似乎没有明显的对应关系, 但一些来自福建、安徽、黑龙江等地的菌株在对生长适宜pH值的要求(偏酸且耐酸)上与它们在培养性状、营养体亲合群以及RAPD聚类结果上表现出了很高的一致性。至于这些地理相距甚远、树种又有所不同的菌株为何会如此相似, 仍有待进一步研究。
值得一提的是, 本研究运用数学模型拟合方法, 根据松杉球壳孢供试菌株在系列温度和系列pH值下培养0 ~ 72 h的连续生长量, 构建出各菌株营养生长的最适温度和最适pH值, 这在国内尚属首次。研究中鉴于松杉球壳孢在PDA平板上(25 ℃, 无光照)生长96 h时, 部分菌株生长已扩至全皿, 故在进行生长与模型拟合数据处理中, 均采用各菌株在各种条件下0 ~ 72h的连续生长量, 这有利于较准确连续地跟踪计测出其最适温度和最适pH值的变化峰值。该研究不仅将各菌株对生长环境的适性差异定量化地展现出来, 而且对进一步深入分析我国各地区松树枯梢病的发生和流行特点有着重要的参考价值。
朗奎健, 唐守正. 1989. IBMPC系列程序集. 北京: 中国林业出版社.
|
钱焕延编.计算方法.上海: 上海交通大学出版社, 1988: 33~ 39
|
吴小芹. 1999. 全球松树枯梢病发生状况与防治策略. 世界林业研究, 12(1): 16-21. |
吴小芹. 2000. 中国松树枯梢病菌营养体亲和性研究. 林业科学, 36(1): 47-52. |
吴小芹. 2000. 松枯梢病菌的培养性状和致病力变异及其相互关系. 南京林业大学学报, 24(2): 16-20. DOI:10.3969/j.issn.1000-2006.2000.02.004 |
吴小芹, 黄敏仁, 尹佟明. 2000. 中国松树枯梢病菌遗传多态性的RAPD分析. 林业科学, 36(4): 32-38. DOI:10.3321/j.issn:1001-7488.2000.04.006 |
Keen A, Snits T F C. 1989. Application of a mathematical function for a temperature optimum curve to establish differences in growth between isolates of a fungus. Netherlands Journal of Plant Pathology, 95(1): 37-49. |