 2002, Vol. 38
2002, Vol. 38文章信息
- 王克勤, 王百田, 王斌瑞, 高海平.
- Wang Keqin, Wang Baitian, Wang Binrui, Gao Haiping.
- 集水造林不同密度林分生长研究
- STUDIES ON THE GROWTH OF FORESTS WITH DIFFERENT DENSITY IN THE SYSTEM OF AFFORESTATION BY WATER-HARVESTING
- 林业科学, 2002, 38(2): 54-60.
- Scientia Silvae Sinicae, 2002, 38(2): 54-60.
-
文章历史
- 收稿日期:2000-08-04
-
作者相关文章


2. 北京林业大学 北京 100083;
3. 山西省方山县林业局 方山 033100
2. Beijing Forestry University Beijing 100083;
3. Bureau of Forestry, Fangshan County, Shanxi Province Fangshan 033100
刺槐(Robinia pseudoacacia)林是黄土高原干旱半干旱区植被恢复的主要人工林类型之一, 对改善这一地区的生态环境、防治水土流失, 乃至对调节黄河流域的水文状况发挥着重要的作用(王佑民等, 1994; 李克亮, 1986)。但现存的大部分人工刺槐林群落密度普遍偏高, 由于水分的不足, 不仅影响到林木个体的继续生长发育(Wang Keqin et al., 1998), 而且造成了严重的人工林地土壤退化现象(王克勤等, 1998)。在年降水量550 mm的陕西安塞地区, 10 a生刺槐林平均胸径仅为7~8 cm, 在年降水570 mm的山西吉县, 16 a生刺槐林平均胸径仅6.32 cm, 在山西河曲, 10 a生刺槐林地0~5 m土层土壤水分含量10月底仅为2.6%~4.0%, 但在年降雨量不足500 mm的山西方山采用集水造林措施, 土壤水分基本满足林分需要, 5 a生刺槐林平均胸径即达7.1 cm(杨维西, 1996)。这说明造林技术对人工林的生长和人工林土壤的影响很大。而以前的人工林营造大多采用常规造林方法, 在造林方法对林木生长的影响研究中, 主要集中在立地条件、种源选择和造林密度等技术措施对人工林生长的影响(鄢志明等, 1999; 温佐吾等, 1998; 陈广太等, 1984), 其中以密度影响林分生长的研究占大多数, 并且得出了各种生长模型(刘财富等, 2000; 刘君然, 1994; 温佐吾等, 2000)。在集水造林系统中, 影响林木生长的水分限制性因子在时空上发生了根本的变化, 以上研究结果不一定适用。而集水造林技术在我国仍是一个“新生事物”, 研究和应用的范围不十分广泛, 尽管有人对采用集水技术后林木生长状况做了一些分析和研究(郑彩霞等, 1995), 但采用何种集水造林模式更有利于林分的生长, 还需要对林分的生长规律进行深入研究, 以期为集水造林的进一步发展和推广应用提供理论依据。
1 试验地概况详见另文(王克勤等, 1998)。
2 材料及方法集水措施营造的15 a生刺槐林, 有840、1110、2220和3330株·hm-2 4种密度, 每株林木有1.5 m2的植树带, 微型集水区表面为自然坡面, 均位于立地条件基本一致的同一坡面, 分别标记为15Y-1、15Y-2、15Y-3和15Y-4。为了比较集水造林与常规造林的差异, 还选择了密度同为1000株·hm-2的9 a生集水造林和常规造林方法营造的刺槐林, 集水造林每株林木有相同面积的植树带(2 m2)和微型集水区(8 m2), 微型集水区表面为压实拍光处理, 常规造林只有植树带(每株林木2 m2), 没有微型集水区, 两种林分位于立地条件基本一致的同一坡面。各林分试验小区面积均为0.1333 hm2。集水造林模式如图 1所示。

|
图 1 集水造林模式图 Fig. 1 The model of afforestation by water-harvesting |
标准雨量计观测林内外降雨量, LNW -50A型中子土壤水分仪测定土壤水分, 标准径流场观测径流量。采用每木调查法测定林木生长量, 标准木法计算林分蓄积量, 标准木解析法确定林分的年龄a及相对应的胸径、树高、材积的连年生长量。由于各密度试验小区林木生长比较均匀, 同径阶标准木数量较多(每一林分不少于15株), 为了不破坏试验地的完整性, 在各林分所有标准木中仅各选1株作为标准解析木, 另选2株用生长锥法分析。在标准解析木全部叶片中抽样, 用LI-3000A叶面积仪测定样叶面积, 用重量折算标准解析木叶面积。
3 结果分析 3.1 林木生长过程 3.1.1 树高生长图 2是15 a生刺槐的树高总生长量、平均生长量和连年生长量的变化曲线。表明密度较小的3种林分, 在7~8 a之前树高生长速度较快, 之后将迅速下降, 平均生长量从7 a时开始下降, 连年生长量最高峰出现在4~6 a, 林分15Y-1、15Y-2和15Y-3的连年生长量最大值分别达到2.48m、1.95 m和1.60 m; 而密度最大的林分15Y-4连年生长量在2~4 a间达到最高峰1.50 m, 在4~5 a时树高生长开始大幅度减缓, 平均生长量从3 a时开始下降。如果以林分15Y-1的连年生长量最大值为100%计, 则其余3种林分分别占到78.63%、64.52%和60.48%, 15Y-1与其他林分之间、15Y-2与15Y-3和15Y-4之间差异显著。各林分在10 a之后树高生长基本处于停滞状态, 接近于各林分树高的最大值, 从小到大4种密度的15 a生刺槐林树高分别达到12.10、10.31、8.45和7.22 m, 如果以林分15Y-1的总生长量为100%计, 则其余3种林分分别占85.21%、69.83%和59.67%。集水造林中随着林分密度的增大树高生长量减小的这种现象与常规造林的研究结果(孙时轩, 1992)有较大的区别, 从后面的植树带汇流量分析中可以看出导致这种结果的原因。

|
图 2 15 a生不同密度刺槐林树高生长变化曲线 Fig. 2 The curves of the height increment of 15 years old black locust forest with different densities
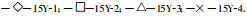
|
上述结果是不同集水造林密度间树高生长的差异。同一密度的集水造林与常规造林之间差异如何?图 3是两者的树高生长过程曲线。树高总生长量的差距随年龄的增长越来越大, 9 a时两者相差15%;平均生长量的变化趋势两者相一致, 但常规造林总是比集水造林低14%左右; 连年生长量的最大值相差不大(分别为1.41 m和1.34 m), 但其他年份仍相差10%~15%, 其中在干旱的1999年两者相差近2.5倍, 常规造林的树高生长量几乎为0。

|
图 3 9 a生同密度集水造林和常规造林刺槐林树高生长变化曲 Fig. 3 The curves of the height increment of 9 years old black locust forest built by water-harvesting and conventional measure with same density

|
林分密度对胸径生长的影响最为显著。本研究4种不同密度的集水造林林分的胸径生长过程(图 4)表明:各林分胸径总生长量在7 a以前增长较快, 但7 a以后随着林分密度的减小增长幅度较大, 以致在15 a时从小到大4种密度林分的胸径总生长量分别达到17.14、11.61、9.17和6.56 cm, 如果以林分15Y-1的胸径总生长量为100%计, 则其余3种林分分别占67.75%、53.51%和38.28%, 各密度林分间相差悬殊; 密度最大的林分15Y-4平均生长量从4~5 a时开始下降, 而其余3种林分则从6~7 a时开始下降; 林分15Y-4的连年生长量最大值出现在2~4 a, 另外3种林分出现在4~6 a, 4种林分的连年生长量最大值也随着林分密度的增大而减小, 分别为2.76、2.01、1.66和1.12 cm, 7 a之后各林分连年生长量有较大幅度下降, 11~12 a时又出现一个较小的生长高峰, 之后迅速下降, 15 a时降到最低点。

|
图 4 15 a生不同密度刺槐林胸径生长变化曲线 Fig. 4 The curves of the DBH increment of 15 years old black locust forest with different densities

|
集水造林与常规造林胸径生长过程(图 5)表明了:集水造林胸径总生长量增长较快, 到9 a时两者相差达28.48%, 分别为9.88 cm和7.69 cm; 2种造林技术平均生长量始终保持着30%~50%的差距; 集水造林的连年生长量在同一年份也比常规造林大, 其中连年生长量最大值出现在4 a, 分别达到1.92 cm和1.33 cm, 到8 a时两者的差距减小。在同种密度下集水造林能促进林木胸径的生长。

|
图 5 9 a生同密度集水造林和常规造林刺槐林胸径生长变化曲线 Fig. 5 The curves of the DBH increment of 9 years old black locust forest built by water-harvesting and conventional measure with same density
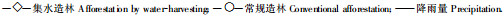
|
材积的大小主要决定于树高、胸径和干型, 由于不同造林密度下各林分的树高和胸径生长差异显著, 因而造成了材积生长的显著差异。15 a生各密度林分的材积生长过程(图 6)表明:材积总生长量在3 a以前增长缓慢, 之后直线上升, 11 a以后又有所减缓, 但密度最小的林分15Y-1增长的速度明显最快, 15 a时密度从小到大各林分材积总生长量分别达到0.2634、0.1030、0.0566和0.0263 m3, 如果以林分15Y-1的材积总生长量为100%计, 则其余3种林分仅占39.10%、21.51%和9.98%;平均生长量在3 a以前增长缓慢, 密度较小的2种林分在11 a以后有下降趋势, 密度较大的2种林分在6 a以后已开始减缓; 连年生长量年际间的变幅较大, 林分密度越小变幅越大, 各林分3 a以前都比较小, 之后增长较快, 在11 a都出现最高峰或次高峰, 12 a之后迅速减小, 但各林分间的差距仍比较大, 林分15Y-3和15Y-4一直处于较低的生长水平。

|
图 6 15 a生不同密度刺槐林材积生长变化曲线 Fig. 6 The curves of the volume increment of 15 years old black locust forest with different densities

|
同密度集水造林和常规造林材积生长过程(图 7)表明: 2种林分材积总生长量、平均生长量和连年生长量的变化趋势极为相近, 但在量的大小上有非常显著的差异。在2 a以前2林分各生长量相差不大, 从3 a时开始拉大差距, 尤其从5 a时差距急剧加大, 9 a时2林分的材积总生长量分别达到0.0647 3m和0.0339 m3, 平均生长量和连年生长量从2 a开始集水造林比常规造林总是大50%~60%。

|
图 7 9 a生同密度集水造林和常规造林刺槐林材积生长变化曲线 Fig. 7 The curves of the volume increment of 9 years old black locust forest built by water-harvesting and conventional measure with same density
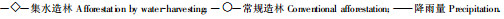
|
林分群体的生产力水平可用林分蓄积量衡量。用标准木法计算各林分的蓄积量, 15 a生从小到大4种密度刺槐林的蓄积量分别为214.58、110.29、121.35和84.03 m3·hm-2, 如果以林分15Y-1的蓄积量为100%计, 则其余3种林分仅占51.40%、56.09%和39.16%, 林分15Y-1的蓄积量大约是林分15Y-2和15Y-3的2倍、林分15Y-4的2.5倍, 林分15Y-2与15Y-3之间差异不显著, 但比15Y-4的蓄积量大30% ~40%。集水造林随着林分密度的增大蓄积量大幅度减小与常规造林及天然林的研究结果(马钦彦, 1983; 刘君然, 1994)有较大的差别, 这主要与水分的分配有关。
通过集水造林与常规造林的对比, 同密度的集水造林林分蓄积量是常规造林蓄积量的2倍, 9 a生集水造林和常规造林的刺槐林蓄积量分别为62.73 m3·hm-2和32.57 m3·hm-2, 采用集水造林技术之后使林分生产力有很大的提高。
3.3 降雨量及植树带径流汇集量(汇流量)与林分生长在干旱半干旱地区影响林木生长的所有环境因子中, 水分是关键性因子(王斌瑞等, 1996; 陈广太等, 1993), 而这些地区林木生长主要依靠天然降水, 降水的缺丰对林木生长的影响将极为显著。山西方山的降雨变率较大, 最大降水量曾达到775.6 mm(1967), 最小降水量只有302.6 mm(1965), 这种降雨特征势必会影响该地区林木生长的特点。但采取集水造林措施之后, 使降雨在时空上得到重新分配, 会使该地区林木原有的生长规律发生一定的改变。在林分营造之后经连续对试验区林内外降雨量和径流观测, 并计算出各林分生长季的植树带汇流量(表 1)。结果表明, 扣除树冠截留量, 林分密度越小, 集水面积越大, 微型集水区产流率越高, 则植树带汇流量越大。林分15Y-1植树带汇流量比林外降雨量高4% ~5%, 林分15Y-2与林外降雨量相当, 而15Y-3和15Y-4则比林外降雨量少10%左右。集水区拍光处理的集水造林植树带汇流量比常规造林多大约30%。植树带汇流量的差异将决定林地植树带土壤水分状况, 从而使林木的生长规律发生变化, 使各林分胸径、树高和材积的总生长量、平均生长量和连年生长量大小随着植树带汇流量的大小产生相应的变化。
|
|
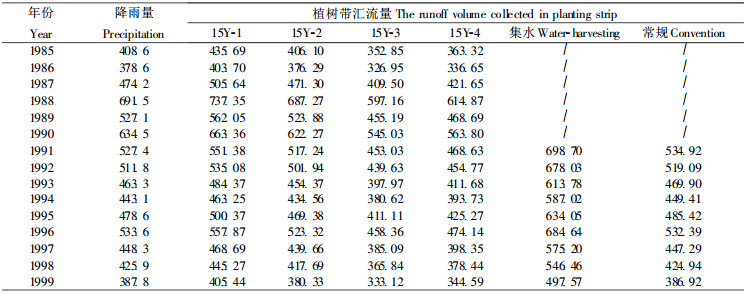
表 1的降雨量变化与上述树高、胸径和材积的连年生长量变化的一致性也充分说明了连年生长量与水分供应量之间的密切关系。降雨量大, 植树带汇流量大, 则林木生长所利用的水量增加, 促进了林木的生长, 反之, 则抑制林木的生长。15 a生各密度林分树高和胸径连年生长量的高峰值出现在4 a或6 a时(1988或1990年), 这2 a的降雨量也是15 a中最高的, 分别为691.5 mm和634.5 mm, 在11~12 a时由于降雨量比年均值高, 胸径和树高的连年生长量又出现了一个小的生长高峰, 使材积连年生长量达到最大值; 在14~15 a时降雨量减少到年均水平以下, 使林木的连年生长量降低到最低点。
9a生集水造林和常规造林刺槐林的连年生长量也与降雨量发生同步变化。从定植后到7 a时, 降雨量普遍偏高(高于年均值), 胸径和树高连年生长量都比较大, 在6~7 a时材积连年生长也达到最大, 8 ~9 a时降雨量减少, 各连年生长量也随之降低, 但集水造林和常规造林之间仍保持着较大的差距。
3.4 土壤含水量与林分生长采取不同的集水面积(林分密度)和集水措施, 使植树带汇流量出现较大的差别, 直接影响到植树带土壤水分状况, 林木将处于不同的土壤水分供给水平。从1996年生长季各林分定期观测可知:不同密度之间和集水造林与常规造林之间, 植树带土壤水分差异较大(王克勤等, 2000)。9 a生2种不同方法营造的刺槐林, 林木平均叶面积相差较大, 蒸腾耗水量不同。1996年集水造林林木平均叶面积为0.2314 m2·tree-1, 年蒸腾耗水1277.89 kg·tree-1, 常规造林这2项值分别为0.1442 m2·tree-1和864.23 kg· tree-1, 集水造林比常规造林单株叶面积和耗水量都大, 而土壤含水量偏高, 更说明了集水造林使植树带汇流量增加, 在一定范围内使林木生长所需要的水分得到保证, 增强了林木的生理代谢功能, 促进林木的生长。在水分亏缺的干旱半干旱区, 林木生长对林地土壤水分状况将有十分敏感的反应。从前面的分析可知, 林木生长量随着林分密度的减小和植树带汇流量的增加而增大, 正是林木对不同土壤水分响应的结果。
4 结论干旱半干旱地区林木的生长主要受水分条件的制约, 水分条件的改善能显著促进林木的生长和提高林分生产水平。
集水造林使植树带汇流量出现较大的差异, 林分密度越小(微型集水区面积越大), 汇流量越大, 林分的生长状况越好。如果以密度最小的林分15Y-1植树带汇流量为100%计, 则林分15Y-2、15Y-3和15Y-4的植树带汇流量分别占93.60%、81.77%和84.43%, 林分15Y-1的年均土壤含水量比林分15Y-3和15Y-4高约20%~25%。由于水分条件的差异和密度效应, 从小到大4种密度林分15 a时树高分别为12.10、10.31、8.45和7.22 m, 胸径分别达到17.14、11.61、9.17和6.56 cm, 林分蓄积量分别为214.58、110.29、121.35和84.03 m3·hm-2, 集水造林各密度林分间生长情况差异显著。随着林分密度的增大, 连年生长量最大值的出现将提前, 生长减缓的时间也较早。集水造林密度大于1000株·hm-2时林分的生长情况较差。
同密度的集水造林和常规造林间植树带汇流量年总量相差约30%左右, 植树带土壤含水率两者相差约10%, 林木处于不同的水分条件下, 造成生长上的差异。9 a时集水造林和常规造林的树高生长分别达到9.13 m和7.04 m, 胸径生长分别达到9.88 cm和7.69 cm, 林分蓄积量分别为62.73 m3·hm-2和32.57 m3·hm-2, 集水造林能促进林木的生长, 提高林分的生产力水平。
集水造林的水分来源是天然降水, 植树带汇流量的年际之差决定于降雨量, 因而林木的连年生长量随着降雨量出现年际变化。15 a生的刺槐林树高和胸径的连年生长量最大值出现在4~6 a, 这时的降雨量也是15 a中最高的, 达到630 mm以上, 在11~12 a时由于降雨量比年均值高, 连年生长量又出现了一个小的生长高峰, 在14~15 a时降雨量减少到年均水平以下, 使林木的连年生长量降低到最低点。9 a生刺槐林的连年生长量也与降雨量发生同步变化, 定植后到7 a时, 降雨量普遍偏高(高于年均值), 胸径和树高连年生长量都比较大, 8~9 a时降雨量减少, 各连年生长量也随之降低。
陈广太, 刘凤山, 王文全. 1984. 太行山刺槐林的调查研究. 河北农学院学报, 9(2): 64-68. |
陈广太, 王文全, 袁玉欣. 1993. 河北花岗岩丘陵地的立地特征与刺槐生长分析. 北京林业大学学报, 15(1): 58-65. |
李克亮. 1986. 中国林业发展战略问题研究. 第一版. 北京: 中国林业出版, 3-21.
|
刘财富, 黄金东, 范俊岗. 2000. 辽宁省刺槐人工林生产力的研究. 林业科学, 36(1): 41-46. DOI:10.3321/j.issn:1001-7488.2000.01.007 |
刘君然. 1994. 林分密度理论及应用. 第一版. 北京: 中国林业出版社, 9-27.
|
马钦彦. 1983. 华北油松人工林单株林木的生物量. 北京林业大学学报, 5(4): 1-16. |
孙时轩. 1992. 造林学. 北京: 中国林业出版, 249-253.
|
王斌瑞, 王百田. 1996. 黄土高原径流林业. 北京: 中国林业出版社, 1-50.
|
王克勤, 王斌瑞. 1998. 集水造林防止人工林植被土壤干化的初步研究. 林业科学, 34(4): 14-21. DOI:10.3321/j.issn:1001-7488.1998.04.003 |
王克勤, 王斌瑞. 2000. 集水造林林分水分生产力研究. 林业科学, 36(sp. 1): 1-9. |
王佐民, 刘秉正. 1994. 黄土高原防护林生态特征. 第一版. 北京: 中国林业出, 105-132.
|
温佐吾, 谢双喜, 周远超, 等. 1998. 不同造林技术措施对马尾松幼林生长影响的研究. 林业科学, 34(6): 39-49. DOI:10.3321/j.issn:1001-7488.1998.06.005 |
温佐吾, 谢双喜, 周远超, 等. 2000. 造林密度对马尾松林分生长、木材造纸特性及经济效益的影响. 林业科学, 36(sp. 1): 36-43. |
鄢志明, 邓健, 张万玉, 等. 1999. 华山松林分生长量的研究. 林业科技通讯, (1): 9-15. |
杨维西. 1996. 试论我国北方地区人工植被的土壤干化问题. 林业科学, 32(1): 78-85. |
郑彩霞, 王斌瑞. 1995. 黄土高原坡面林地不同集水措施对林木生长的影响. 北京林业大学学报, 17(4): 56-61. |
Wang Keqin, Yang Xiaohui. 1998. The Optimum Thinning Intensity and Water Productivity of Black Locust Forest on Loess Plateau. Journal of Beijing Forestry University (in English), 7(2): 19-27. |