 2002, Vol. 38
2002, Vol. 38文章信息
- 何斌, 温远光, 袁霞, 梁宏温, 刘世荣.
- He Bin, Wen Yuanguang, Yuan Xia, Liang Hongwen, Liu Shirong.
- 广西英罗港不同红树植物群落土壤理化性质与酶活性的研究
- STUDIES ON SOIL PHYSICAL AND CHEMICAL PROPERTIES AND ENZYME ACTIVITIES OF DIFFERENT MANGROVE COMMUNITIES IN YINGLUO BAY OF GUANGXI
- 林业科学, 2002, 38(2): 21-26.
- Scientia Silvae Sinicae, 2002, 38(2): 21-26.
-
文章历史
- 收稿日期:2000-10-31
-
作者相关文章


2. 中国林业科学研究院森林生态环境与保护研究所 北京 100091
2. Institute of Forest ecology, Environment and Protection, CAF Beijing 100091
红树植物群落(或红树林)是生长在热带、亚热带海岸潮滩盐渍土的特有的木本植物群落(林鹏, 1997), 具有防浪护岸固沙, 保护和改善当地生态系统, 促进滩涂养殖业发展等重要作用。土壤基质是影响红树植物群落的生态分布并得以维持的重要原因, 有关土壤理化性质对不同红树植物分布和生长发育的影响, 近年来已有不少报道(杨萍如等, 1987; 温肇穆, 1987; 张希然等, 1991; 蓝福生等, 1994), 而作为土壤肥力标志之一的土壤酶活性, 在农业土壤和陆地森林土壤中已进行了大量研究(Zantua, et al., 1977; 黄世伟, 1981; 关松荫等, 1984; Peruic et al., 1984; 张其水等, 1990; 潘映华等, 1995), 对于处于海陆交界地带间的红树植物群落土壤酶活性也已有报道(张银龙等, 1999)。本文对广西英罗港不同红树植物群落土壤理化性质和酶活性及其相互关系进行较系统的研究, 其目的是为红树植物的保护、恢复和管理提供科学依据。
1 研究区的自然概况研究地点位于广西山口国家级红树林海洋生态自然保护区英罗港红树植物群落核心区内, 东经109°43″, 北纬21°28″, 属北热带季风区。年平均气温22.4℃, 极端最高气温37.4℃, 极端最低气温-0.8℃。雨量较充沛, 年平均降雨量为1816.5 mm, 雨量多集中在5~9月。年平均相对湿度为81.8% (温远光, 1999)。英罗港红树植物群落面积约80 hm2, 属港湾红树植物群落类型, 群落类型主要有白骨壤(Avicennia marina)群落、桐花树(Aegiceras corniculatum)群落、秋茄(Kandelia candel)群落、红海榄(Rhizophora stylosa)群落和木榄(Bruguiera gymnorrhiza)群落等群落, 并依次由外滩向内滩呈带状分布, 其间存在着一些过渡的群落类型, 反映了群落的演替进程。
2 研究方法 2.1 样品采集通过对群落的全面踏查, 以群落组成和滩位为主导因子, 分别在5种不同红树植物群落内各设置3个10 m×5 m的样方, 采用径级平均木法并增加权重因素测算群落地上部分的生物量(温远光, 1999), 用取土器在样方内按表层(0~20 cm)和第二层(20~40 cm) 3点采集土壤样品, 把相同样方同一层次土壤等比例混合, 风干过筛后备用。采样时间为1997-07。
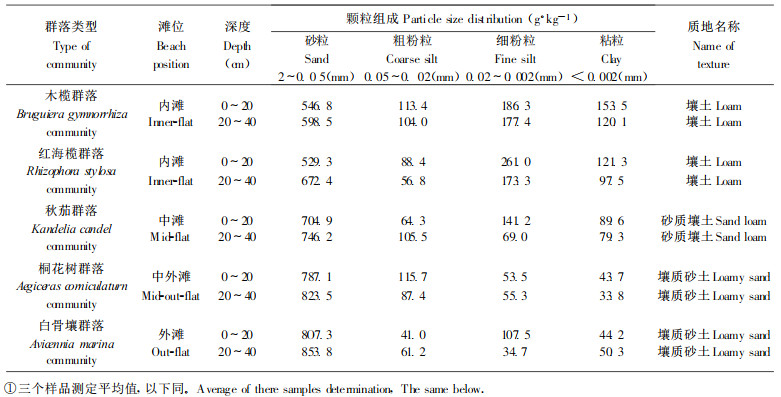
2.2 分析方法 2.2.1 土壤机械组成用吸管法测定, 并按美国土壤质地分类法进行土壤质地命名。
2.2.2 土壤化学性质pH值用电位法, 全氮用硫酸-高氯酸消化-氨电极法(何斌, 1992), 有机质、水解氮、全磷、速效磷和盐分含量按常规方法测定(中国科学院南京土壤研究所, 1978)。
2.2.3 土壤酶活性过氧化氢酶用J.C.Tohonson和K.L.Temple法; 多酚氧化酶用K.A.Kosol法; 蔗糖酶用T.A.Ⅲ epóakoba法; 蛋白酶用G.Hoffmann和K.Teicher法; 脲酶用G.Hoffmann与K.Teicher法; 酸性磷酸酶用G.Hoffmann法(Ф.X.哈兹耶夫, 1980; 严昶升, 1988)。
3 结果与分析 3.1 英罗港不同红树植物群落土壤机械组成英罗港5种红树植物群落土壤机械组成分析结果(见表 1)表明, 各群落土壤砂粒(2~0.05 mm)含量在529.3~853.8 g·kg-1之间, 无论是表层(0~20 cm)土壤还是第二层(20~40 cm)土壤, 其土壤颗粒含量的变化均为砂粒 > 粉粒(0.05~0.002 mm) > 粘粒(< 0.002 mm), 这与我国其它地方多数红树林潮滩土的土壤颗粒组成相一致(张希然等, 1991)。从水平分布看, 土壤中砂粒含量为白骨壤群落 > 桐花树群落 > 秋茄群落 > 红海榄群落 > 木榄群落, 即从内滩到外滩, 砂粒含量逐渐升高, 粉粒和粘粒含量则呈现相反趋势, 从而导致了不同群落土壤质地的差异, 内滩木榄群落和红海榄群落为壤土, 中滩的秋茄群落为砂质壤土, 中外滩的桐花树群落和外滩的白骨壤群落则均为壤质砂土, 土壤质地越来越不适于红树植物的生长; 而从垂直分布看, 土壤中砂粒含量为表层 < 第二层, 粉粒、粘粒含量多数是表层 > 第二层。这说明潮汐、波浪等环境因子对不同滩位土壤颗粒组成影响的差异, 更重要的是反映了不同红树植物对生态环境和土壤基质的要求和适应, 以及红树植物的改善土壤物理性状和促淤聚泥作用。
|
|
由于红树植物对海水硫酸根的选择性吸收, 致使其体内含硫量特别高, 深刻影响着土壤发生及其性质, 使土壤具有含硫层, 含硫层中的可氧化硫化物经氧化产生硫酸, 加上红树植物富含单宁, 其残体可分解出单宁酸, 从而导致土壤显著酸化(龚子同, 1994; 林鹏, 1997)。从分析结果(表 2)看, 英罗港5种红树植物群落土壤pH值在2.85~4.12之间, 属强酸性潮滩土, 并低于我国其它地区的多数红树林土枹(张希然等, 1991); 不同群落土壤pH值的变化为白骨壤群落 > 桐花树群落 > 秋茄群落 > 木榄群落 > 红海榄群落, 且呈表层土壤 > 第二层土壤(白骨壤群落除外)的趋势。显然, 这除了与不同红树植物含硫量的差异和群落生物量以及植物残体埋藏层深浅有关外, 也可能与潮水对不同滩位红树植物群落土壤的影响有关。
|
|

潮滩作为一种生境, 由于不同滩位所受到的潮汐、波浪等环境条件影响的差异, 其土壤质地有所不同, 影响着土壤养分和盐分的分配, 导致不同滩位土壤养分和盐分含量存在一定的差异, 从而影响着红树植物的生态分布, 同时又因为不同红树植物的生长速度、残落物量及残落物营养元素含量的不同, 影响了土壤养分的归还和提高, 更加大了各滩位土壤养分水平的差异。从表 2可以看到, 土壤有机质、全氮、全磷、水解氮、速效磷5种养分与盐分含量均为木榄群落 > 红海榄群落 > 秋茄群落 > 桐花树群落 > 白骨壤群落, 这与5种红树植物地上部分的营养水平恰好相反(表 3), 反映了不同红树植物对土壤养分与盐分的要求和适应性, 影响着红树植物群落的演替进程。白骨壤个体较小又最耐贫瘠, 根系特别发达, 所以能成片分布于受波浪、潮汐冲击最强, 土壤质地和肥力水平较差的外滩; 秋茄、桐花树对土壤养分的要求和耐盐性略高于白骨壤, 故主要分布于中滩和中外滩; 内滩的土壤质地良好(壤土), 肥力水平较高(有机质含量50 g·kg-1以上), 受波浪和潮汐的冲击较弱, 所以木榄和红海榄得以在英罗港大量分布, 其中红海榄群落成为我国大陆热带海滩面积最大, 发育也较典型的红海榄群落。由此进一步说明不同红树植物群落成带状分布并得以维持除决定于波浪和海潮的影响外, 与土壤条件有着非常密切的关系。
|
|
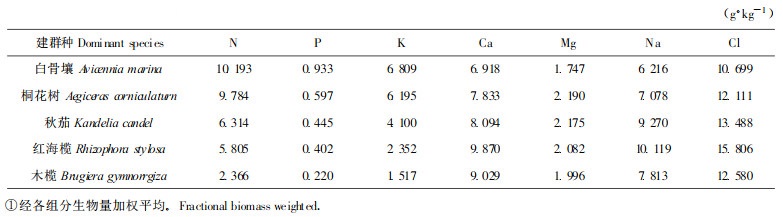
从表 2中还可以看出, 各群落表层土壤的养分含量均大于第二层土壤, 这与多数陆地土壤养分在剖面中的垂直分布相一致, 而盐分含量表现出第二层土壤大于表层土壤, 与我国其它地区多数红树植物土枹呈现相反的趋势(张希然等, 1991; 林景亮, 1990), 这可能与英罗港土壤质地较粗, 砂粒含量较高, 盐分离子容易淋洗下移有关。此外, 英罗港5种红树植物群落土壤的C/N较大, 达24.8~41.0, 明显大于一般陆地土壤, 说明厌氧和强酸条件下对土壤的腐殖化过程产生了较大的阻碍作用。
3.3 英罗港不同红树植物群落土壤酶活性土壤酶是土壤生物化学反应的催化剂, 它参与土壤中物质的转化过程与循环, 促进有机质的分解, 反映土壤生物活性的大小, 因此可以作为土壤肥力的标志之一。表 4表明, 英罗港不同红树植物群落土壤酶活性均存在较明显差异, 蔗糖酶、蛋白酶、脲酶和磷酸酶4种水解酶与氧化还原酶中的过氧化氢酶的酶活性, 其滩位变化规律均呈现内滩 > 中滩 > 外滩, 这与张银龙等(1999)报道的基本一致, 群落变化规律则为红海榄群落 > 木榄群落 > 秋茄群落 > 桐花树群落 > 白骨壤群落, 与各群落地上部分生物量大小变化规律相一致(温远光, 1999), 体现出土壤酶活性与植物现存量的关系, 即单位面积的植物产量越多, 土壤酶活性越高(Pancholy et al.; 张银龙等, 1999); 但多酚氧化酶的变化规律则与此相反。在土壤酶活性的垂直分布中, 各群落土壤的4种水解酶多数为表层土壤 > 第二层土壤, 多酚氧化酶分布不很规则, 与陆地土壤相一致(关松荫等, 1984), 过氧化氢酶活性的垂直分布则呈现相反的规律。由此可见, 不同红树植物群落土壤间和土壤上下层之间的土壤有机残体分解强度以及土壤中腐殖质合成强度均存在一定的差异。
|
|
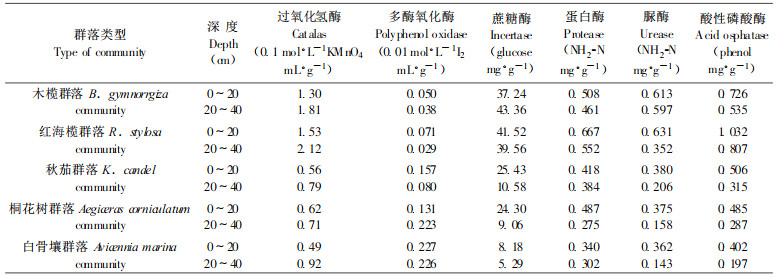
不同红树植物群落土壤的4种水解酶活性均以蔗糖酶大于蛋白酶、脲酶和酸性磷酸酶。可见蔗糖酶是最活跃的水解酶, 说明红树植物群落土壤中有机物的代谢以蔗糖酶最为剧烈, 蛋白酶、脲酶和酸性磷酸酶的代谢相对较弱, 这表明蔗糖酶活性比其它土壤酶活性更能反映土壤的生物学活性强度和土壤熟化程度。
回归分析表明(表 5), 英罗港红树植物群落土壤蔗糖酶、蛋白酶、脲酶和酸性磷酸酶4种水解酶活性相互间呈极显著的线性正相关, 相关系数分别为0.8663、0.8586、0.8584、0.7991、0.9540和0.7949, 过氧化氢酶和多酚氧化酶2种氧化还原酶之间也呈显著正相关, 相关系数为0.7420, 这表明在水解酶之间及氧化还原酶之间各存在相互促进作用, 对环境也有相似的适应性。此外, 在土壤水解酶与氧化还原酶之间, 大多数存在显著的正相关关系, 表明土壤4种水解酶和2种氧化还原酶在促进土壤有机质的转化和参与土壤物质转化和能量交换中, 不仅显示其专有特性, 同时存在着共性关系。
|
|

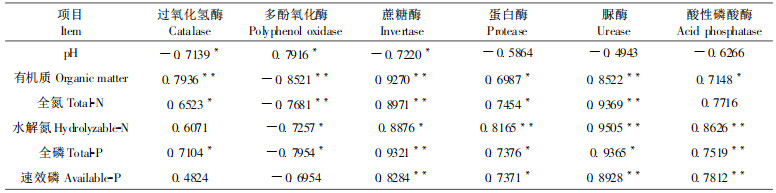
大量研究表明, 农业土壤和陆地森林土壤酶活性与土壤肥力因素有良好的相关性(黄世伟, 1981; 关松荫等, 1984; Peruci et al., 1984)。经过对英罗港红树植物群落土壤酶活性与土壤主要肥力因素进行相关分析, 结果表明(表 6), 4种水解酶活性与土壤有机质、全氮、水解氮、全磷和速效磷均呈极显著或显著正相关(P < 0.01或P < 0.05), 其中蔗糖酶和脲酶与上述5项肥力因素均达到极显著水平。氧化还原酶类中的过氧化氢酶与有机质、全氮、全磷也有显著的正相关关系, 表明土壤酶活性在很大程度上反映了该土壤的营养水平。值得注意的是, 土壤有机质与4种水解酶和过氧化氢酶的活性均存在极显著或显著正相关, 说明土壤有机质不但是其它养分的重要来源和储藏场所, 同时也是土壤酶活性的来源和储藏基地。此外, 土壤pH值与5种土壤酶活性(多酚氧化酶除外)均呈负相关, 其中与过氧化氢酶、蔗糖酶达到显著水平, 而多酚氧化酶与土壤5种养分含量均呈极显著或显著负相关, 这与陆地土壤明显不同, 这可能是红树植物发育下的酸性盐滩土与陆地土壤具有不同性质所致。
|
|
英罗港红树植物群落土壤颗粒组成含量分布为砂粒(2~0.05 mm) > 粉粒(0.05~0.002 mm) > 粘粒(> 0.002 mm), 体现了大多数潮滩土壤的特点。从内滩的木榄群落到中滩的秋茄群落再到外滩的白骨壤群落, 土壤中砂粒含量逐渐增多, 粉粒和粘粒含量逐渐减少, 土壤质地类型由壤土经砂质壤土过渡到壤质砂土。
由于受到土壤质地和红树植物的生物累积与循环作用的不同影响, 英罗港5种红树植物群落的土壤养分和盐分含量均存在明显差异, 表现为木榄群落 > 红海榄群落 > 秋茄群落 > 桐花树群落 > 白骨壤群落, 反映了不同红树植物对土壤养分和盐分的要求和适应性, 表明不同红树植物群落成带状分布并得以维持既决定于潮水的影响, 同时又与土壤条件有密切的关系。
英罗港红树植物群落土壤酶活性, 随所在滩位的不同呈现有规律的梯度变化, 除多酚氧化酶外, 蔗糖酶、蛋白酶、脲酶、酸性磷酸酶4种水解酶与氧化还原酶中的过氧化氢酶活性大小顺序均为红海榄群落 > 木榄群落 > 秋茄群落 > 桐花树群落 > 白骨壤群落, 这与各群落的地上部分生物量变化规律相一致, 它们的相关性均达到极显著或显著正相关(P < 0.01或P < 0.05), 如表层土壤蔗糖酶活性与各群落的生物现存量的相关系数为0.9286**, 第二层土壤为0.8528**。可见红树林的存在, 对土壤酶活性的提高有明显的促进作用。
英罗港红树植物群落土壤酶活性与土壤肥力因素有密切的关系, 其中土壤蔗糖酶、蛋白酶、脲酶、酸性磷酸酶4种水解酶与土壤有机质、全氮、水解氮、全磷、速效磷5种肥力因素均存在极显著或显著的正相关, 氧化还原酶中的多酚氧化酶则与之呈显著的负相关。水解酶活性之间、氧化还原酶活性之间、以及水解酶与氧化还原酶之间也存在一定程度的相关关系。由此可见, 与陆地土壤一样, 土壤酶活性也可作为评价酸性盐滩土土壤肥力的指标之一。
龚子同, 张效朴. 1994. 中国的红树林与酸性硫酸盐土. 土壤学报, 31(1): 86-93. DOI:10.3321/j.issn:0564-3929.1994.01.014 |
关松荫, 沈桂琴, 孟昭鹏, 等. 1984. 我国主要土壤剖面酶活性状况. 土壤学报, 21(4): 368-381. |
哈兹耶夫ФХ.土壤酶活性.北京: 科学出版社, 1980
|
何斌. 1992. 植物氮磷钾的自控远红外快速联合消化和测定法. 理化检验(化学分册), 28(4): 44-45. |
黄世伟. 1981. 土壤酶活性与土壤肥力. 土壤通报, (4): 37-39. |
蓝福生, 李瑞棠, 陈平, 等. 1994. 广西海滩红树林与土壤的关系. 广西植物, 14(1): 54-59. |
林鹏. 1997. 中国红树林生态系. 北京: 科学出版社.
|
潘映华, 施亚琴, 李振高.红壤区不同植被下土壤酶活性的研究.见: 中国科学院红壤生态实验站编, 红壤生态系统研究(第三集).北京: 中国农业科技出版社, 1995, 172~178
|
温远光. 1999. 广西英罗港5种红树植物的生物量和生产力. 广西科学, 6(2): 142-14. DOI:10.3969/j.issn.1005-9164.1999.02.019 |
温肇穆. 1987. 广西红树林植物化学元素含量的初步研究. 热带亚热带林业科技, (2): 9-24. |
严昶升. 1987. 土壤肥力研究方法. 北京: 农业出版社.
|
杨萍如, 何金海, 刘滕辉. 1987. 红树林及其土壤. 自然资源学报, 2(1): 32-37. DOI:10.3321/j.issn:1000-3037.1987.01.004 |
张其水, 俞新妥. 1990. 杉木林地土壤酶活性的分布特征研究. 福建林学院学报, 10(4): 377-381. |
张希然, 罗旋, 陈研华. 1991. 红树林和酸性潮滩土. 自然资源学报, 6(1): 55-62. DOI:10.3321/j.issn:1000-3037.1991.01.006 |
张银龙, 林鹏. 1999. 秋茄红树林土壤酶活性时空动态. 厦门大学学报(自然科学版), 38(1): 129-136. DOI:10.3321/j.issn:0438-0479.1999.01.025 |
中国科学院南京土壤研究所编.土壤理化分析.上海: 上海科学技术出版社, 1978
|
Pancholy S K, Rich E L. Soil enzymes in relation to old field succession: Amylase, cellulase, invertase, dehydrogenase andurease.Soil Sci Soc Amer Proc, 1973, 37: 47~50
|
Peruci P, Scarponi L, Businelli M. 1984. Enzyme activities in clay loam soil amended with various crop residue. Plant and Soil, 81: 345-351. DOI:10.1007/BF02323049 |
Zantua M I, Dumeil I C, Bramner J B. 1977. Relationships between soilurease activity and other properties. SoilSci Soc Amer J, 41: 350-352. DOI:10.2136/sssaj1977.03615995004100020036x |