 2002, Vol. 38
2002, Vol. 38文章信息
- 秦建华, Stephen J. Titus, Shongming Huang.
- Qin Jianhua, Stephen J. Titus, Shongming Huang.
- 林分生长与产量模型系统研究综述
- TEH STUDY SYNTHESIS OF STAND GROWTH AND YIELD MODEL SYSTEMS
- 林业科学, 2002, 38(1): 122-129.
- Scientia Silvae Sinicae, 2002, 38(1): 122-129.
-
文章历史
- 收稿日期:2000-01-24
-
作者相关文章


2. Dept. of Renwable Resources, University of Alberta, Edmonton, Alberta, Canada, T6G 2H1;
3. Alberta Environment Protection, Edmonton, Alberta, Canada, T5K 2M4
2. Dept. of Renwable Resources, University of Alberta, Edmonton, Alberta, Canada, T6G 2H1;
3. Alberta Environment Protection, Edmonton, Alberta, Canada, T5K 2M4
材分生长与产量预测模型系统应用于森林经营管理, 可提高林业生产经营水平, 应用于教学, 可显著提高教学效果。国外学者20世纪70年代开始研制林分生长与产量预测模型系统, 经过30 a的发展, 目前已进入成熟阶段, 正向着估测精度更高、更符合生产实际的系统模型方向发展。本文旨在综述目前国外林分生长与产量预测模型系统的研究现状和发展趋势。
1 影响林分生长和产量的主要因子以前林学家认为影响林分生长和产量的主要因子是立地质量(立地指数)、林分年龄及林分密度。其中立地指数影响最大。Assmann(1955)研究了挪威云杉, Kennl (1973)山毛榉, Schmidt (1973) Sots松, Schutz和Badoux (1979)栎树, 发现对于给定的立地指数和其他条件相同的情况下, 林分产量变化较大。Bavaria和Bradley (1966)将这种现象归因于“产量水平”。Sterba和Monserud (1993)研究发现, 相同立地指数的不同生境类型的林分间产量差异较大。林分的生长和产量与树种特性密切相关。生境类型是林分树种组成、树种间相互作用及树种与环境相互作用的结果。Sterba和Monserud (1995)研究花旗松混交林产量时, 用数量化(0-1)方法将林分树种组成引入林分最大胸高断面积方程。目前, 关于生境类型对林分产量影响的研究不多, 特别是针对树种间相互作用及树种与环境间相互作用对林分产量的影响的研究, 几乎未见报道。许多学者是划分生境类型研制林分生长和产量预测模型系统。
生态系统的结构决定其功能。林分结构影响环境资源(如光、生长空间、营养元素等)的利用, 从而影响林分生长、死亡和产量。Sterba和Monserud (1993)认为林分产量不仅是林分静态属性如立地指数和生境类型的响应特征, 林分的动态属性—林分结构—也强烈地影响林分产量, 特别是异龄混交林。秦建华等(1995)研究发现, 不同疏伐方法改变了林分结构, 对杉木人工林生长、竞争和产量产生显著影响。Sterba和Monserud (1993)认为不同林分间“产量水平”的差异主要归因于林分的生境类型和林分结构。因此, 影响林分产量的主要因子应是立地指数(SI)、林分年龄(A)、林分密度(N)、林分生境类型(HT)和林分结构(SS)。即林分产量(Y林分)是上述5个因子的函数。

|
(1) |
林分结构包括林分直径分布、空间分布、林木年龄结构等。由于直径分布可部分反映空间利用, 如直径分布宽度可反映林冠长度和林分垂直空间利用。直径分布越宽, 林冠长度越长, 垂直空间利用越充分。直径分布形状可反映林分生长历史和竞争死亡情况。Buongiorno和Dahir (1994)认为直径分布是林分结构的最佳表征。Weibull、SB、β、指数分布等分布函数常被用来描述林分直径分布。其中Weibull分布以其灵活性和广泛适用性而被普遍采用。Stiff (1979)用左截的Weibull分布函数描述混交林结构。Hyink (1980)用左截的Weibull函数描述异龄林结构。Murphy和Farrar (1982)发现异龄林的直径分布呈反J形, 用指数分布函数更好。Sterba和Monserud (1993)提出用D1.31.5分布的偏斜度描述林分结构。即D1.31.5为0时, 林分产量最高。异龄林D1.31.5分布的偏斜度增加, 接近于2, 林分产量显著下降。
2 林分生长和产量预测模型系统的分类200年前欧洲学者就编制了林分产量表, 根据林分年龄预测林分产量。Buckman (1962)研制了林分生长和产量模型。20世纪70年代随着计算机的普及, 林学家开始研究林分生长和产量预测模型系统, 80年代后期趋于成熟。林分生长和产量预测模型系统是林分生长和产量的模拟器或生成器。是在广泛收集林分生长数据基础上, 应用竞争密度理论、自然稀疏规律等, 分析、建立各种经验性关系、生长模型、死亡模型等模型, 确定林分的统计平均产量。林分生长和产量预测模型系统包括树高生长模型、直径生长模型、林木死亡模型、树冠动态模型、疏伐或施肥等干预模型、林木进阶模型等子模型。其核心是生长模型和死亡模型。不同模型系统间的主要差别是生长模型的建模思路和途径的不同。依之可分为林分水平的、径级水平的和单株木水平的3类林分生长和产量预测模型系统。
2.1 林分水平的林分生长和产量预测模型系统该类模型系统中的生长、死亡等模型是以林分水平的变量, 如林分年龄、立地指数、林分密度等作自变量。生长、死亡等模型都是提供林分的平均特征。应用Chapman-Rechards方程、Logistic方程、多元回归方程等, 建立林分优势树高和胸高断面积预测方程。应用林分形数方程(Y林分=F林分H林分G林分)或二元材积表预测林分产量。林分直径分布可以推测或假定(常为正态分布)。该类模型系统提供的单株林木的信息很少, 林业生产上的应用受到限制。Curtis等(1981)研制的DFSIM (Douglas-Fir Stand Simulator)即为此类模型系统。该系统应用竞争密度理论和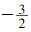
该类模型系统是以林木直径级(林木组)为生长和产量预测的基本单元。林分产量表、转移矩阵模型系统、林分直径分布模型系统等属此类型。林分产量表和转移矩阵模型系统允许部分林木在一些生长期转移几个直径级, 易造成林分产量的过高估计。因此, 更窄直径级的直径分布模型系统发展较多、影响较大。直径分布模型系统用概率密度函数(pdf)描述林分直径分布。以立地指数和林分年龄为自变量, 用Chapman-Rechards方程估测各径级的平均树高。用材积方程计算各径级材积, 累计可得全林分材积。该类模型系统能够提供林分直径分布(结构)详细信息, 突出了林分结构对林分生长和产量的影响。Feduccia等(1979), Bailey和Dell (1973), Hyink和Moser (1983), Lenhart (1988)等研制了直径分布的林分生长和产量预测模型系统。通常用来描述林分直径分布的概率密度函数为Weibull函数。Matney和Sullivan (1982), Hyink和Moser (1983), Burk和Burkhart (1984), Mantey (1987)等研制了概率密度函数参数的恢复程序。可用林分水平变量, 如总产量、总断面积、林分密度、立地指数、林分年龄、最小直径、算术平均直径、平方平均直径等, 来恢复林分直径概率分布函数的参数。以此来研究林分直径分布结构的变化动态。
2.3 单株木水平的林分生长和产量预测模型系统单株木水平的林分生长和产量预测模型系统是以典型木特征为基础, 典型木特征即代表径级平均木特征, 生长和死亡等模型应用于典型木, 获得典型木特征, 以此推导出径级和全林分的生长和产量。也称之为林木特征清单模型系统。该类模型系统是以单株木树高生长、直径生长为基本生长单元, 用林木当前大小和林分水平的特征作自变量, 建立树高和直径的生长增量方程, 结合林分直径分布模型、死亡模型、树冠动态模型等形成的模型系统。该类模型系统能概括单株木和林分两级水平的特征。详细的单株木特征包括树种、胸径、树高、树冠大小、树冠比率等。Munro (1974)依据是否与单株林木的位置有关, 划分为与距离无关的和与距离有关的两种单株木水平的林分生长和产量预测模型系统。
Mitchell和Cameron (1985)研制的TASS (Tree And Stand Simulator)即为与距离有关的单株木水平的林分生长和产量预测模型系统。该模型系统借助于最大生长、当前大小、树冠比率和竞争指数, 应用修改的Chapman-Rechards方程模拟林木断面积和树高生长。林木最大生长是立地质量、年龄和空旷地生长树木大小的函数。该模型系统给每株林木分配了一个协调的位置, 突出了林分生长与林木间竞争的关系, 可以获得最详细的单株木信息。但由于采用了与林木位置有关的竞争指数变量, 限制了其应用的普遍性和灵活性。Stage (1973), Wykoff (1986;1990)研制的Prognosis (Stand Prognosis Model)和Titus (1992)研制的MGM (Mixedwood Growth Model)等模型系统, 属于与距离无关的单株木水平的林分生长和产量预测模型系统。该类模型系统灵活性很强, 可以引入干扰模型、进阶模型等, 允许使用者调控许多参数, 可提供确定性或统计性操作。可以为广范围立地和生长条件下的林分, 提供较可靠的林分生长和产量预测。该类模型系统可以提供单株林木和林分2方面信息, 而且可以灵活地加入施肥、疏伐等干扰子模型, 教学和生产上的应用价值较高, 是目前影响最大的模型系统。Prognosis 1986年已推出了第五版, 已被推广应用到12个地区。它还具有“自身校准”功能, 可以用临时标准地的生长增量, 修正系统中生长增量方程的参数, 使模型系统适合于当地的林分。
3 林分生长和产量预测模型系统的主要模型林分生长和产量预测模型系统包括树高生长模型、直径生长模型、死亡模型、直径分布模型、树冠动态模型、干扰模型、进阶模型等。其核心是生长模型和死亡模型。生长模型可用Gompertz方程、Chapman-Rechards、Logistic、Bertalanffy、Korf等方程拟合。最常用的是Chapman-Rechards方程和Logistic方程。
3.1 树高生长模型林分生长和产量预测模型系统中采用的树高生长模型有两种。一是树高与立地指数和年龄间的关系方程; 二是树高增量方程。林分水平和径级水平的模型系统用前者, 单株木水平的模型系统用后者。
3.1.1 树高生长方程Payandeh等(1974), Monserud (1984), Newnhan (1988)等学者应用了5参数的Chapman-Rechards方程模拟林分树高生长、立地指数曲线。该方程应用很普遍。
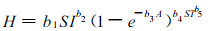
|
(2) |
其中, H为林分优势树高或共优势树高, b1, b2, b3, b4, b5为回归参数。
Logistic方程也常被用来模拟林分优势树高生长。Monserud (1984)用方程(3)模拟花旗松的优势树高生长。
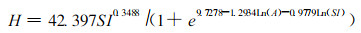
|
(3) |
另外, Ek-Payandeh方程、多项式回归方程等也可用来描述林分优势树高生长。
3.1.2 树高增量方程单株木水平的模型系统均是以树高增量方程为基础。树高生长增量随年龄增加而增加, 达一定年龄后随年龄增加而下降, 呈单峰状态。不同研究者采用不同的方程模拟树高生长增量的变化。如Huang (1992)应用SAS中二阶段非线性回归程序, 用方程(4)模拟美洲山杨的树高生长增量(ΔH)。

|
(4) |
其中, ΔH为某生长期树高增量, BA为生长期开始时林分胸高断面积, D1.3为生长期开始时林木的胸径, ΔD为生长期林木直径增量, SC=该树种总胸高断面积/林分全部林木总胸高断面积, 反映该树种在林分中的地位, SPI为立地生产力指数, 是指胸高直径达到20 cm时的树高值, b1, b2, b3, b4, b5, b6为回归参数。
3.2 直径或胸高断面积生长模型 3.2.1 林分水平的胸高断面积生长或增量模型该类方程均以林分水平的变量, 如林分密度、年龄等, 作自变量, 预测林分胸高断面积或胸高断面积增量。如Pienaar和Shiver (1986), Pienaar和Rheney (1993)用方程(5)预测林分胸高断面积。
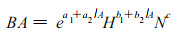
|
(5) |
其中, H为林分平均优势树高, a1, a2, b1, b2, c为回归参数。
Nelson (1963), Vanclay (1988)等研制了林分胸高断面积增量方程。Nelson (1963)认为同龄林分的胸高断面积生长增量, 随林分年龄增加而逐渐下降, 随立地指数增加而增加, 随林分胸高断面积增加而下降。他用方程(6)描述林分胸高断面积增量(ΔBA)的变化。
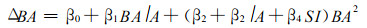
|
(6) |
其中, β0, β1, β2, β3, β4为回归参数。
3.2.2 单株木水平的直径或胸高断面积增量模型该类方程是以影响林木直径生长的因子, 如年龄、立地指数、林分树种组成、林木在林分中地位等, 作自变量, 建立单株木的直径增量方程或胸高断面积增量方程。West (1980), Shifley (1987)研究认为直径增量方程或胸高断面积增量方程的估测精度没有什么差别。如Quicke和Meldahl (1994)用方程(7)模拟长叶松单株木胸高断面积年增量(BAI)。

|
(7) |
其中, A为林分优势木和共优势木平均年龄, BAL为大于目标林木的所有林木胸高断面积之和, a0, a1, b0, c0, c1, c2为回归参数。
Yao (1997)用方程(8)估测白云杉单株木的直径增量。
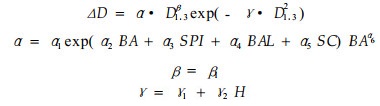
|
(8) |
其中, ΔD为某生长期的直径增量, D1.3为生长期开始时的林木胸径, H为林木树高, α1, α2, α3, α4, α5, α6, β1, γ1, γ2为回归参数。
建立生长模型的另一种途径是以空旷地生长树木的生长量为最大生长量, 乘上一系列修正因子, 即得具体林木的生长量。如Amaties等(1989)研制的单株木直径增量方程(9), 将最大直径增量乘上竞争性修正因子和林木在林分中地位修正因子。
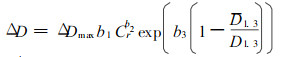
|
(9) |
其中, Cr为树冠比率(即树冠长度/树高), D1.3为林分平方平均直径, ΔDmax为空旷地生长树木的直径增量, b1, b2, b3为回归参数。
3.3 林分的死亡估测模型林分产量预测不仅需要准确的生长估计, 也需要准确的死亡估计。林木死亡受多方面因子影响。如环境因子、生理因子、病虫害因子、自然因子(风、雪、动物)等。这些因子又与林分立地、年龄、密度等相互关联, 导致林木死亡具有很大的偶然性和难以预测性。因此林分死亡估测是最困难的、可靠性最低的。Stage和Renner (1988)发现温带森林估测产量变动的80%是死亡估计的不准确造成的。林分死亡估测模型分为单株木水平的死亡估测模型和林分水平的死亡估测模型两种。
3.3.1 单株木水平的死亡估测模型Monserud (1976), Hamilton (1980), Yao (1996), Zhang (1997)等绝大多数学者采用Logistic方程估测林木的存活概率或死亡概率。一些反映林木特征和林分特征的因子被用作估测自变量。如林木直径增量、树冠比率、林木竞争指数、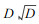
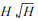
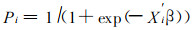
|
(10) |
其中, Pi为第i株林木的存活概率, Xi′为多个变量组成的向量的转置。β为回归系数向量。
从单株木存活概率, 推断林木是否死亡, 还涉及林木死亡判断标准的确定。而该标准随年龄、立地质量等而变化。且推测全林分死亡估计, 存在累计误差。单株木水平的死亡估测模型中包含多个变量, 收集数据难度大, 应用受到一定限制。
3.3.2 林分水平的死亡估测模型林分水平的死亡估测模型, 是用林分水平的变量建立回归方程或死亡概率分布模型。Bailey (1980)假设林分中的死亡林木正比于直径分布, 这显然是不符合实际的。因为小树比大树更容易因竞争而死亡。Ford (1975)认为人工林中死亡林木呈规则分布。Kent和Dress (1979;1980)认为原初随机分布的林分, 死亡木保持随机分布。规则的人工林, 随着年龄增加, 死亡林木趋于随机分布。尽管死亡发生于生长期全过程, Cao (1997)假设全生长期的死亡只发生在生长期开始时, 从而简化了林分水平的死亡估测模型。他将直径分布的Weibull函数乘上林分死亡修正函数, 得到新的直径分布函数。总之, 不管是单株木水平的死亡估测模型, 还是林分水平的死亡估测模型, 均未达到可靠的估测精度。对于年龄较高林分, 单株木水平的林分死亡估测往往偏低。可以应用竞争密度理论和
一些相关模型集合构成模型系统。系统模型是从整体出发, 充分考虑系统组成成份间的相互联系和相互作用, 以系统结构为基础, 建立一系列相互协调模型的集合。模型系统中子模型是单独拟合的。而系统模型则是建立联立方程系统, 用同一数据同时拟合所有方程的回归参数。Pindyck和Rubinfeld (1981)将相关方程系统分为3类。即表面上不相关的方程系统、递归方程系统(11)和联立方程系统(12)。
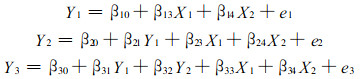
|
(11) |
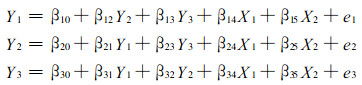
|
(12) |
表面上不相关的方程系统, 一般用多元回归程序拟合。但如果方程间误差项相关, 则方程间存在交叉相关, 应该用Zellner (1962)的3步拟合程序拟合。递归方程系统。如果方程间误差项不相关, 则用普通最小二乘法(OLS)拟合。否则, 不是严格的递归方程系统, 而应建立联立方程系统。联立方程系统, 则需用同时回归拟合技术-SAS软件系统中的二阶段最小二乘法(2SLS)或三阶段最小二乘法(3SLS)程序进行拟合。
林木是一级水平的生态系统, 树高生长、直径生长、树冠生长、死亡间都是密切相关的。最明显、最密切的是树高生长和直径生长、直径生长和死亡。Clutter (1963)提出应建立相互协调的林分生长和产量预测模型。Furnival和Wilson (1971), Murphy和Steritzke (1979), Murphy和Beltz (1981), Borders和Bailey (1986), Gregoire (1987), Judge (1988)等学者都认为单株木水平的生长模型的回归系数间存在逻辑约束关系。所有变量的观测值来自于同一林木个体, 变量间是同时相关的。单株木水平的模型系统中方程间是交叉相关的, 应建立联立方程系统, 用2SLS或3SLS同时拟合所有方程的回归参数。习惯上, 许多学者研究单株木水平的模型系统时, 生长模型和死亡模型均是用OLS单独拟合的。尽管不少学者考虑到了方程间相关。如Stage (1975), Huang (1992)在树高增量方程中, 用直径增量作预测变量。Hamilton (1986), Yao (1996)等在死亡模型中用直径增量为预测变量。但他们都是用同一数据库用OLS对每个方程单独拟合的, 未考虑林木系统的同时性。Hasenauar等(1998)认为忽视了生长模型系统的同时性特性, 方程间的参数估计不相容的、有偏的, 导致模型系统估测精度的累计损失。Zhang等(1997)认为联立方程系统提供的单株木直径增量估计与非联立方程系统的相同, 但可提供更准确的直径级和全林分的断面积估计。Hasenauar等比较了用OLS单独拟合和用了3SLS同时拟合树高增量方程、断面积增量方程等, 发现用3SLS同时拟合的方程系统, 可提供更准确的模型形式(变量构成)。用OLS拟合的模型中两个重要变量, 在用3SLS拟合的模型中并不重要, 可删去, 体现了变量间的同时相关性。Borders等(1986), Borders (1989), Huang (1992), Zhang等(1997), Hasenauer等(1998)对该方面进行了探讨, 不同程度地提高了所建模型的精度。
林分是另一级水平的生态系统。生态系统的结构决定其功能。系统模型的第2个特征是强调林分结构对林分产量的影响。如前所述, 影响林分产量的主要因子是立地指数、年龄、密度、生境类型和林分结构。Monserud (1984)认为树高是立地指数、年龄和生境类型的函数。直径体现了林分密度的影响。许多林分生长和产量预测模型系统都是针对特定生境类型研制的。目前模型系统中欠考虑的是林分结构对林分产量的影响。该方面研究仅局限于用直径分布反映林分结构, 不够充分、准确。未考虑环境因子、树种组成、干扰因子、经营措施等对林分结构的影响。因此, 系统模型应将结构向量引进生长模型、死亡模型等模型, 以提高系统模型的真实性和估测精度。这方面的研究有待于进一步的探索。
5 生物学模型的建模步骤 5.1 概念模型(Conceptual Models)概念模型是科学过程的延伸, 也可以说是科学过程本身的体现。在整个系统的框架内, 将系统中可操作的成分、相互作用、发生机理与研究目标、研究对象相结合, 进行统一考虑、总结, 形成概念模型。该阶段应分析系统组成成分间的内在关系和作用机理, 明确系统的主导功能关系(forcing functions)和驱动变量(driving variables)。选择、确定模型的类型、目标、功能、可操作性(实用性)。并对模型的通用性、真实性、准确性等方面有所评估或预计。构思概念模型是建模的最重要的一步, 直接影响所建模型的质量和水平。

|
图 1 生物学模型的建模步骤流程图 Fig. 1 Flow diagram of the steps for developing biological models |
图解模型是将概念模型用图形、文字、数学符号具体体现和表达。该阶段主要解决的问题是模型的复杂性。真实系统非常复杂、多变量、多子系统。应用大系统理论, 通过聚合多过程, 将系统中相类似的组成成分归为功能组, 划分子系统, 达到系统简化。系统中不少变量所反映的系统组成成分、关系是交叉的或部分重叠的, 通过筛选最能反映系统状态、组成成分间相互作用、系统主导功能关系的变量, 用于建立模型。该阶段应明确子目标, 划分、确定子系统及其主导功能关系和驱动变量。
5.3 数学模型(Mathematical Models)将系统中主导功能关系、状态变量间关系及状态变量的行为表现, 用数学公式(模型)表达。该阶段务必选准适合的数学公式, 对系统的主导功能关系及主要状态变量进行描述, 必须明确不同状态变量的初始值、改变状态变量的主导功能关系(forcing functions)、状态变量间相关关系。选择、确定的数学模型最好是理论性模型(Theoretical Models), 如D2H可推广应用到许多地区, 否则, 模型的应用范围只能局限于研制模型所用数据的收集地区。
5.4 计算机模型(Computer Models)选择适合的参数拟合程序, 将数学模型翻译成计算机模型(用计算机语言), 应用所收集数据进行试拟, 根据对模型的敏感性分析, 细致调整模型的变量数量和形式, 最终将子模型集成系统整体模型。该阶段要解决模型精度与模型复杂性问题, 确定最终模型形式。敏感性分析是确定模型中变量及其系数对模型输出项影响的重要程度。通过测试给同一变量的不同输入值, 比较模型输出项变化大小, 分析该变量的敏感性。模型系统的模拟引擎(Simulation Engine)可以选用C语言等多种语言编写, 操作界面可用excel, 使用者易学易用。
5.5 模型的检验(Validation of Models)检验模型模拟真实世界的吻合程度。要求检验是独立检验。一般在研制模型前将数据随机划分为2部分, 2/3数据用于拟合模型参数, 1/3数据用于模型的检验。通常应用下列途径对模型进行检验: (1)比较和模拟(Comparison and Simulation)。给模型和更大的系统输入相同的输入项, 检查2者输出结果是否相同。模拟是将自然系统的实测值与模型的模拟值相比较, 检验其吻合度。如用前述方法保留的1/3数据来检验模型。(2)专家检验。将模型递送给专家进行评估、检验。对于模型系统, 先对子模型逐个检验, 然后检验模型系统整体, 找出影响较大、误差较大的子模型, 进行修正。如对于林分生长和产量预测模型系统, 可用传统的、较可靠的林分产量表进行模型系统的整体检验。
6 讨论林分水平、径级水平和单株木水平的林分生长和产量预测模型系统的着重点不同。林分水平的模型系统注重的是林分整体状态, 并不关心单株林木的发展过程和林木间的相互作用。致力于提高模型系统的通用性和整体估测精度。单株木水平的模型系统着眼于林木间相互作用及林木生长和死亡的过程描述, 致力于提高模型系统的真实性和准确性。径级水平的模型系统突出了林分结构对林分生长和产量的影响。Levins(1996)指出生物学模型不可能同时达到通用性、真实性和准确性3个目标, 只能达到其中2个。理想的选择是提高林分生长和产量预测的真实性和准确性。只有与距离无关的单株木水平的林分生长和产量预测模型系统的改进余地、发展潜力最大。林木和林分是两级不同水平的生态系统。应该从生态系统特性着手, 注重生态系统的整体性、同时性、结构决定性, 充分考虑系统组成成份间的相互联系和相互作用。不仅要考虑单株木树高生长、直径生长和死亡间的交叉相关, 而且也要考虑林木间的相互作用, 兼顾林分整体估计的协调性。突出林分结构对林分生长和产量的影响, 将结构向量引进生长和死亡估测模型, 建立树高生长、直径生长、树冠生长、死亡等的联立方程系统, 应该用2SLS或3SLS同时拟合所有方程的回归参数, 从而获得相互协调的林分生长和产量预测的系统化模型。提高林分生长和产量预测的真实性和准确性。
秦建华, 阮益初, 姜志林, 等. 1995. 不同疏伐方法对杉木人工林结构、生长和竞争的影响. 浙江林学院学报, 12(4): 360-366. |
唐守正, 李希菲, 等. 1993. 林分生长模型研究的进展. 林业科学研究, 6(6): 672-679. DOI:10.3321/j.issn:1001-1498.1993.06.018 |
Amaro A, Reed D, et al. 1998. Modeling dominant height growth: Ecualyptus plantations in Portugal. For Sci, 44(1): 37-46. |
Amateis R L, Burkhart H E, et al. 1989. Diameter increment and survival equations for loblolly pine trees growing in thinned and unthinned plantations on cutover, site prepared lands. South J Appl For, 13(4): 170-174. DOI:10.1093/sjaf/13.4.170 |
Borders B E. 1989. System of equations in forest stand modeling. For Sci, 35(2): 548-556. |
Borders B E, Bailey R L. 1986. A compatible system of growth and yield equations for slash pine fitted with restricted three-stage least squares. For Sci, 32(1): 185-201. |
Cao Q V. 1997. A method to distribute mortality in diameter distribution models. For Sci, 43(3): 435-442. |
Daniels R F, Burkhart H E. 1988. An integrated system of forest stand models. For Ecol Manage, 23: 159-177. DOI:10.1016/0378-1127(88)90080-1 |
Dolphy K L. A diameter increment model for red fir in Califormia and southern Oregon. USDA For Serv Res Pap PSW, 1992
|
Gregorie T G. 1987. Generalized error structure for forestry yield models. For Sci, 33(2): 423-444. |
Hall C A S and Day J W, Jr. Ecosystem modeling in theory and practice : An introduction with case histories. John Wiley & Sons, 1977
|
Hamilton D A. 1986. Jr. A logistic model of mortality in thinned and unthinned mixed conifer stands of Northern Idaho. For Sci, 32(4): 989-1000. |
Hasenauer H, Monserud R A, et al. 1998. Using simultaneous regression techniques with individual tree growth models. For Sci, 44(1): 87-95. |
Huang S. Nonlinear simultaneous diameter and height growth models for major Alberta tree species. PhD thesis. University of Alberta, 1992
|
Hyink D M, Moser J W. 1983. Jr. A generalized framework for projecting forest yield and stand structure using diameter distribution. For Sci, 29(1): 85-95. |
Lenhart J D. 1988. Diameter distribution yield-prediction system for unthinned loblolly and slash pine plantations on non-old-fields in east Texas. South J Appl For, 12(4): 239-242. DOI:10.1093/sjaf/12.4.239 |
Monserud R A. 1984. Height growth and site index curves for inland Douglas-fir based on stem analysis data and forest habitat. For Sci, 30(4): 943-965. |
Monserud R A, Sterba H. 1996. A basal increment model for individual trees growing in even and uneven-aged forest stands in Austria. For Ecol Manage, 80: 57-80. DOI:10.1016/0378-1127(95)03638-5 |
Murphy P A, Farrar R M. 1988. Jr. A framework for stand structure projection of uneven-aged loblolly-shortleaf pine stands. For Sci, 34(2): 321-332. |
O'Hara K L, Oliver C D. 1988. Three-dimensional representation of Douglas-fir volume growth: comparison of growth and yield models with stand data. For Sci, 34(2): 321-332. |
Pienaar L V, Shiver B D. 1986. Basal area prediction and projection equations for pine plantations. For Sci, 32(3): 626-633. |
Pienaar L V, Rheney J W. 1995. Modeling stand level growth and yield response to silvicultural treatments. For Sci, 41(3): 629-638. |
Quicke H E, Meldahl R S, et al. 1994. Basal area growth of individual tree: a model derived from a regional longleaf pine growth study. For Sci, 40(3): 528-542. |
Sterba H, Monserud R A. 1995. Potential volume yield for mixed-species Douglas-fir stands in the Northern Rocky Mountains. For Sci, 41(3): 531-545. |
Sterba H, Monserud R A. 1993. The maximum density concept applied to uneven-aged mixed-species stands. For Sci, 39(3): 432-452. |
Vanclay J K. 1995. Growth models for tropical forests: a synthesis of models and methods. For Sci, 41(1): 7-42. |
Wykoff W R. 1990. A basal increment model for individual conifers in the Northern Rocky Mountains. For Sci, 36(4): 1077-1104. |
Wykoff W R, Crookston N L, and Stage A R. User' s guide to the stand prognosis model. USDA For Serv, GTR INT-133, 1982
|
Yao X. Modeling juvenile growth and mortality relationships in mixedwood stands of Alberta. PhD thesis. University of Alberta, 1992
|
Zeide B. 1993. Analysis of growth equations. For Sci, 39(3): 594-616. |
Zhang S, Amateis R L, Burkhart H E. 1997. Constraining individual tree diameter increment and survival models for loblolly pine plantations. For Sci, 43(3): 414-423. |