 2002, Vol. 38
2002, Vol. 38文章信息
- 殷亚方, 姜笑梅, 魏令波.
- Yin Yafang, Jiang Xiaomei, Wei Lingbo.
- 毛白杨形成层的活动周期及其POD同工酶的变化
- PERIODICITY OF CAMBIUM ACTIVITY AND CHANGES OF PEROXIDASE ISOZYMES IN POPULUS TOMENTOSA CARR.
- 林业科学, 2002, 38(1): 103-110.
- Scientia Silvae Sinicae, 2002, 38(1): 103-110.
-
文章历史
- 收稿日期:2000-10-11
-
作者相关文章


2. 中国科学院发育生物学研究所 北京 100080
2. Research Institute of Developmental Biology, CAS Beijing 100080
树木的次生维管系统(secondary vascular system, SVS)是由次生韧皮部、维管形成层和次生木质部组成, 它是一个动态的四维发育系统(Chaffey, 1999)。通过形成层原始细胞的分裂和分化产生次生韧皮部和次生木质部(Fahn, 1982)。温带树木的维管形成层活动具有明显的周期性, 但不同树种形成层的活动式样存在较大差异(Fahn, 1982; 张英伯等, 1982; 崔克明等, 1992; 1993; 1995; Luo et al., 1995; 张仲鸣等, 1997)。形成层活动过程中发生的生理生化变化, 是研究形成层活动机理的一个重要方面。树木次生木质部管状分子的分化是一种典型的细胞程序化死亡(PCD)方式, 在植物发育和细胞分化的不同阶段, 编码同工酶基因的表达不同(Roberts et al., 1988), 同工酶对细胞的生理调节功能也不同。目前有关同工酶在树木形成层活动以及其衍生木质部管状分子的分化过程中的作用机理仍不清楚(Reinecke et al., 1987; 王雅清等, 1998; 1999)。尽管已有关于POD同工酶在植物形成层活动过程中变化的报道, 但多是集中在形成层从休眠期向活动期的转化过程(崔克明等, 1993; Luo et al., 1995)。杨树是我国北方重要的造林树种, 同时在阔叶树种SVS发育的研究中, 杨属树种作为新的模式树种正受到日益广泛的关注, 对其形成层的活动深入开展研究, 可为木材形成的分子生物学研究奠定细胞生物学基础, 有助于了解木材的形成机理并为木材性质的生物改良提供理论依据。目前有关杨树形成层活动及其与同工酶变化之间关系的研究还未见报道。本研究通过组织解剖学方法, 运用聚丙烯酰胺凝胶电泳(PAGE)技术, 在分析毛白杨整个年活动周期POD同工酶酶带的基础上, 结合了组织化学定位方法, 对毛白杨形成层的活动式样以及整个年活动周期中POD同工酶的变化对次生组织分化形成的影响进行有益的探讨。
1 材料与方法 1.1 材料在位于北京的中国林业科学研究院院内选择树龄约为30a的雄性人工林毛白杨。取材时间1999- 03-15 ~ 2000-03-23。1999-03-15 ~ 04-28以及1999-08-04 ~ 10-19, 每隔1周取材1次, 其它时间每隔1个月取材1次。每次取材时选择3株树, 每株树各取2根长约20 cm的3 ~ 4 a生健康枝条, 每批共计6根, 立即放入-20 ℃冰箱中保存。
1.2 方法 1.2.1 解剖学及组织化学观察从每批采集的6根枝条上分别取下0.5 cm×0.5 cm×1 cm的组织块, 将其等分成2块, 均包括韧皮部、形成层和木质部。其中1块用FAA固定后, 聚已二醇渗透包埋, 在滑走切片机上切片, 厚度为15 ~ 20 μm, 番红染色后拍照, 再用Olympus AH-2型显微镜观察, 参照崔克明等(1992)描述的方法, 每张切片统计15个径向细胞列; 另1块用滑走切片机切新鲜切片, 厚度为20μm, 利用联苯胺通过过氧化氢经酶促作用脱氢而产生蓝色络合物, 对POD进行组织化学定位, 方法参照《植物生理学实验手册》(薛应龙主编, 1985), 染色后立即用Olympus AH-2型显微镜拍照。
1.2.2 取样及电泳方法在每批材料中随机取出3根枝条, 截成10 cm长。参照崔克明等(1993)的方法, 每根枝条用蒸馏水冲洗3次, 并剥去树皮, 轻轻刮取形成层区域的植物组织。刮取的组织混匀后, 随机称取200 mg, 加1.5 mL的提取液(40 %蔗糖溶液), 冰浴条件下迅速研磨, 10 kr.min-1离心10 min, 取上清液。采用聚丙烯酰胺凝胶垂直板状不连续电泳技术, Tris-甘氨酸(pH 8.3)电极缓冲系统, 分离胶浓度12.5 %, pH 8.8;隔层胶浓度4 %, pH 6.8。取50 μL组织上清液加入样品槽底部, 溴酚兰作指示剂, 在4 ℃下进行电泳, 始电流为25 mA, 始电压为220V, 当指示剂移动到下层电泳液面以下1 cm处, 停止电泳。电泳约4 h, 终电压为550V, 凝胶制备及POD同工酶的染色参照胡能书的方法(胡能书等, 1985), 染色时间严格控制在3 min.
1.2.3 电泳结果分析方法根据酶带染色结果绘制酶带类型图, 并拍下照片, 再利用岛津CS-930色谱扫描仪在540 nm波长下对毛白杨形成层POD同工酶带进行扫描, 得到各酶带的相对酶量。
2 实验结果 2.1 不同时期的形成层活动状态毛白杨形成层的年活动周期中, 形成层带的变化以及木质部和韧皮部细胞的产生, 同其物候特点的变化有着密切的关系。
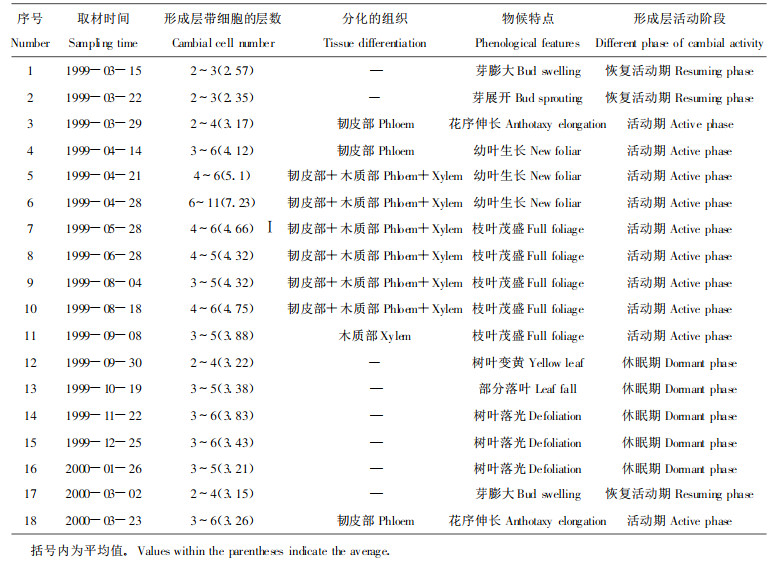
2.1.1 形成层带的变化1999-03-15, 在春季芽展开前一周形成层细胞就已经具备了不同于休眠期的特点(表 1)。取材的形成层带有2 ~ 3层具有恢复活动期特点的细胞, 细胞径向直径增大, 细胞壁特别是弦壁变薄, 细胞排列比较整齐(图版Ⅰ-1), 这一阶段形成层带细胞层数无明显变化(图 1-A, P > 0.05)。4月中旬, 随着幼叶的生长, 形成层细胞层数开始显著增加(图 1-A, P < 0.05), 到4月底形成层处于活动旺盛期, 细胞层数达到6 ~ 11层, 由于细胞分裂频繁, 此时细胞的排列较不规则(图版Ⅰ-4)。至6月底又恢复到4月中旬的水平。8月中旬, 形成层细胞的径壁开始加厚(图版Ⅰ-5), 随后一直到活动期结束, 形成层带细胞层数没有大的变化(图 1-A)。进入休眠期后, 形成层细胞具有典型的休眠期特点, 细胞形状扁平且径壁变厚, 细胞内含物丰富(图版Ⅰ-7), 细胞层数基本保持不变, 直到翌年春天形成层开始恢复活动。
|
|

|
图版Ⅰ Plate Ⅰ |

|
图 1 不同组织细胞层数的季节变化 Fig. 1 Seasonal changes of the number of various tissue cell layers |
1999年3月底, 在芽展开后一周开始出现正在扩大的未成熟韧皮部细胞(图版Ⅰ-2), 较木质部早约20 d (表 1), 到4月28日达到最大值(3-5层), 随后保持一个较稳定的水平, 直到9月8日停止分化未成熟韧皮部细胞(图 1-A)。4月28日出现成熟的韧皮部细胞, 由此可见, 韧皮部细胞的分化成熟约需30天。韧皮部细胞总数一直缓慢地增加, 到8月18日以后就不再明显增加(图 1-B, P >0.05)。
2.1.3 木质部细胞的产生1999年3月底和4月中旬, 韧皮部细胞已经开始产生, 但直到4月21日幼叶生长后一周, 才出现2层未成熟木质部细胞(包括正在扩大的导管和木纤维)(表 1)(图版Ⅰ-3), 5月28日未成熟木质部细胞层数迅速增加(图 1-A, P < 0.05), 6月28日达到最大值, 随后急剧减少(P < 0.01), 8月中旬分化的未成熟木质部和韧皮部细胞胞壁(图版Ⅰ-5)明显较前期(图版Ⅰ-3, 4)要厚, 到9月30日未见到未成熟木质部细胞(图版Ⅰ-6), 表明此时形成层已停止向木质部分化细胞。5月底开始出现多层次生壁加厚的成熟木质部细胞, 可以推测木质部细胞的成熟需要30 ~ 40 d, 从5月28日到8月初, 成熟木质部细胞不断显著增加(图 1-B, P < 0.01), 从木质部细胞总数来看, 增长趋势一直保持到9月30日树叶开始变黄为止(表 1)。
2.2 形成层活动的不同时期POD同工酶酶带的变化由于形成层在不同时期活动状态的变化, 剥皮的难易程度各不相同, 每次取样都尽量在紧贴树皮内侧刮取形成层区域的植物组织, 并使各层组织充分混匀, 以保证各次取样的可对照性。
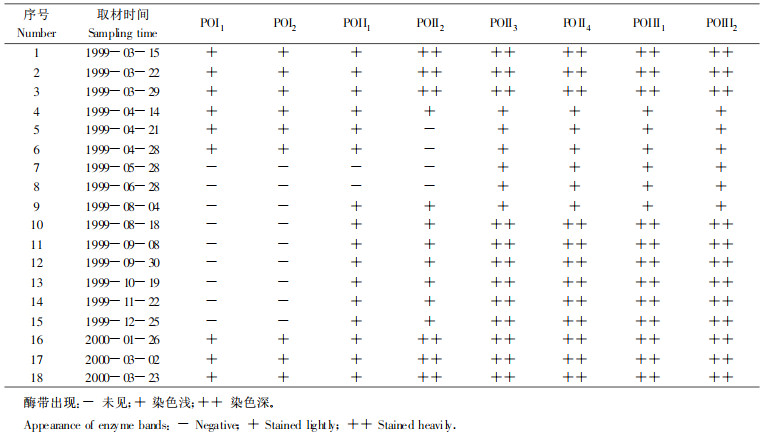
休眠期形成层区域POD同工酶酶带最多, 主要有8条, 分布在3个酶活性区, 根据迁移速度可分为3组:POⅠ, POⅡ, POⅢ。其中:POⅠ包含POⅠ1和POⅠ2; POⅡ包含POⅡ1, POⅡ2, POⅡ3, POⅡ4; POⅢ包含POⅢ1, POⅢ2(表 2; 图 2)。为了方便对酶谱的分析, 各不同时期的POD同工酶酶谱均以此为标准来描述和比较。
|
|

|
图 2 不同时期形成层区域的POD同工酶酶谱及其扫描图 Fig. 2 Zymogram of peroxidase isozyme from electrophoresis and its scanning spectra from the cambial region of Populus tomentosa Carr.twigss A.1999-03-15;B.04-14;C.06-28;D.2000-01-26 Samples collected on March 15, 1999(A), April 14, (B), June 28, (C), January 26, 2000(D) |
通过对不同时期形成层区域组织POD同工酶酶谱的比较分析(表 2)可知:在1999年3月底形成层完全恢复活动之前, POD同工酶酶谱基本相似, 其中POⅡ2, POⅡ3, POⅡ4, POⅢ1, POⅢ2的活性较强, 从4月14日开始, POⅡ2, POⅡ1, POⅡ4, POⅢ1, POⅢ2的活性普遍降低(图 2-B), 到5月28日, POⅠ1, POⅠ2和POⅡ1这3条酶带也消失, 一直持续到8月4日, POⅡ1, POⅡ2又重新出现, 而POⅠ组酶带直到2000年1月26日才又重新出现(图 2-D)。各组酶带在2000年1月26日的活性都与前一年形成层恢复活动之前的酶带一致。
2.3 形成层活动的不同时期POD的组织化学定位观察经组织化学定位表明, POD主要分布在细胞壁及其邻近区域, 染色后形成蓝色络合物, 但因反应时间的长短, 颜色略有差异。POD的分布及活性明显随形成层活动周期发生变化。
当形成层处于休眠期时, 形成层细胞和韧皮部细胞以及部分髓部细胞的POD活性较高, 并且POD在整个形成层区域分布较均匀。韧皮部细胞比形成层细胞的POD活性要略高, 而与形成层细胞紧邻的成熟的木质部细胞POD消失了(图版Ⅱ-1, 2)。
随着形成层的恢复活动, POD的分布也发生变化。在形成层带中, 射线原始细胞比纺锤状原始细胞的POD活性明显要高; 在木质部中, 刚刚分化形成的导管分子和木纤维也有活性较高的POD, 而去年分化的次生壁已加厚的木质部细胞则无POD; 在韧皮部, 除了韧皮纤维团, 无论是刚分化还是成熟的细胞都有分布(图版Ⅱ-3, 4)。
3 讨论 3.1 毛白杨形成层活动的式样生长在温带地区树木形成层的活动具有明显的周期性。大多数松柏类和散孔材双子叶树木的形成层是从膨大展开的芽基部自上而下开始活动(Fahn, 1982)。毛白杨属于散孔材树种, 在春季芽展开前一周形成层细胞就已经具备了不同于休眠期的特点, 说明形成层已经进入了恢复活动期。在芽展开后一周形成层细胞开始分裂, 细胞层数也有所增加, 同时分化出未成熟韧皮部细胞(表 1), 这与散孔材中的构树(Broussonetia papyrifera (L.)Vent.)(崔克明等, 1993)及松柏类的欧洲赤松(Pinus sylvestris Linn.)(崔克明等, 1992)和白皮松(Pinus bungeana Zucc.)(张仲鸣等, 1997)的形成层在芽展开前就开始活动明显不同, 与杜仲(Eucommia ulmoides Oliv.)(Luo et al., 1995)也不完全相同, 其形成层活动刚开始并不象构树那样减少细胞层数, 而是从活动伊始就持续增加。这可能与毛白杨形成层反应的一个显著特点有关, 即形成层活动开始并不象构树那样同时形成韧皮部和木质部, 而是先分化韧皮部, 后分化木质部, 这就能保证在形成层活动刚开始细胞分裂频率较低的情况下, 使得新分裂产生的子细胞数能够略多于分化成韧皮部的细胞数。同时韧皮部筛分子的较早分化可能有利于树木在春季迅速生长时所需营养物质的运输和储藏物质的重新分配(Evert, 1963)。Sachs(1981)提出的生长素流理论认为由芽和幼叶产生的呈波状的IAA流, 沿着形成层带和其周围分化中的维管组织向基运输(Little, 1981; Wodzicki et al., 1987), 从而控制形成层细胞的形态、分裂和排列。毛白杨形成层细胞的首次分裂发生在芽展开之后约一周, 这在一定程度上证实了以上观点。
对于毛白杨, 越冬后未成熟韧皮部细胞的出现比未成熟木质部要早3周左右, 这与张英伯等(1982)的研究结果基本一致。比较未成熟韧皮部和未成熟木质部细胞层数在整个活动期中的变化过程可知:未成熟韧皮部细胞的数量在形成层反应初期快速增长(3月29日到4月21日), 随后是一个持续而缓慢的连续过程, 一直到韧皮部停止分化前20 d, 才逐渐减少; 未成熟的木质部细胞层数在反应初期变化不大, 到了5月底到6月底之间, 增长速度明显高于前期, 随后便急剧下降。
值得注意的是, 与其他研究(崔克明等, 1993; 张仲鸣等, 1997; 罗立新等, 1995; Luo et al., 1995)不同, 毛白杨在4月底到6月底这段时间, 形成层细胞层数迅速下降, 未成熟韧皮部细胞也略有减少, 而未成熟和成熟的木质部细胞则大量增加, 这可能是由于此时正值幼叶的快速生长期, 大量新合成的IAA不断向下运输, 高浓度IAA的存在诱导了未成熟木质部细胞的大量产生及其向成熟木质部细胞的分化, 但并未促进未成熟韧皮部细胞的产生; 同时由于新分化的木质部细胞数量远远大于形成层细胞新分裂所产生的子细胞数, 导致形成层细胞层数迅速减少。崔克明等(1992)认为, IAA对韧皮部的影响主要是促进这一分化过程, 而不影响它的细胞分裂, IAA对木质部产生的促进(李正理等, 1984; Sundberg et al., 1991), 则明显包括产生它的细胞分裂, 也包括其后的分化过程。本研究的结果为IAA分别在树木韧皮部和木质部分化过程中起着不同的作用提供了新的证据。
3.2 POD同工酶的变化与组织分化的关系大多数树木次生组织的分化活动都与形成层区域的IAA水平变化有关(Fahn, 1990)。POD在植物细胞、组织和器官的发生、分化成熟过程中具有重要作用, POD参与细胞壁木质化过程, 影响细胞发育类型和形态大小, 并通过对IAA侧链的氧化调节植物内源IAA的浓度, 从而影响形成层的活动(Reinecke et al., 1987)。
与杜仲(Eucommia ulmoides Oliv.)(Luo et al., 1995)和构树(Broussonetia papyrifera (L.)Vent.)(崔克明等, 1993)相似, POD同工酶活性的的变化与形成层的活动周期有相关关系, 即在活动期其活性低、酶带少, 而恢复活动期和休眠期其活性高且酶带多。
POⅡ2酶带在未成熟木质部细胞出现前一周开始减弱, 4月下旬在POⅡ2酶带消失的同时, 未成熟木质部细胞出现, 随后POⅡ1也消失, 到8月4日这两条酶带又重新出现, 而此时未成熟木质部细胞的数量已急剧减少。POⅡ1和POⅡ2酶带的全部消失, 正值未成熟和成熟的木质部细胞大量增加, 这可能是由于上述时间内IAA侧链氧化酶的活性降低, 提高了内源IAA的浓度, 从而促进了木质部细胞的分化。同时也说明POⅡ1和POⅡ2可能参与了IAA浓度的调节。
POⅠ组酶带在5月底开始消失, 这时形成层细胞层数迅速下降, POⅠ组酶带直到第二年1月份才重新出现, 这表明POⅠ可能通过调节IAA的水平, 激活形成层细胞内的某些酶或合成新的酶, 通过一系列信号传递, 最后使特定的基因类群有序表达(Libbenga et al., 1998), 从而控制了形成层细胞形态、排列和分裂的控制; POⅠ在休眠期当中的重新出现, 是否与毛白杨形成层由生理休眠期(rest)向被动休眠期(quiescence)转化有关, 还需要进一步研究。
POⅡ3、POⅡ4以及POⅢ组酶带在整个形成层活动周期中都存在, 只是活动期活性有所降低, 因此它们可能与IAA浓度的调节无关。
另外通过组织化学观察, 在活动期, 毛白杨形成层纺锤状原始细胞中POD的活性明显比射线原始细胞要小(图版Ⅱ-4), 这可能由于纺锤状原始细胞在活动旺盛期, 为了不断向次生组织产生大量的新细胞, 需要减少横向分裂以避免失去平周分裂能力, 而低活性的POD提高了IAA的浓度, 从而抑制了这种横向分裂的发生(Savidge et al., 1984; Cui et al., 1995)。同时, 在新生导管和木质部纤维尚未加厚的细胞壁中存在着一定活性的POD, 而对于次生壁已加厚的成熟的导管和木质部纤维, POD已经消失, 这表明POD参与了未成熟木质部细胞的分化过程, 至于其具体的作用方式和机理, 还有待于进一步研究。
图版说明
Explanation of Plate Ⅰ
图版Ⅰ毛白杨枝条的部分横切面普通光学显微照片.C.形成层带P.韧皮部X.木质部IP.未成熟韧皮部Ⅸ.未成熟木质部.1.3月15日, 形成层处于恢复活动期, 示形成层细胞内含物明显减少(细箭头), ×120;2.3月29日, 形成层开始活动, 出现未成熟韧皮部细胞, 示正在扩大的筛管分子(细箭头), ×120;3.4月21日, 出现未成熟木质部细胞, 示正在扩大的导管分子(细箭头), ×400;4.4月28日, 形成层带及未成熟木质部细胞层数明显增加, 示正在扩大的导管分子(细箭头), ×400;5.8月18日, 示未成熟木质部细胞(细箭头)和韧皮部细胞(箭头), ×400;6.9月30日, 未成熟木质部和韧皮部细胞已停止分化, ×400;7.12月25日, 形成层处于休眠期, 示形成层细胞内含物丰富(细箭头), ×400。
Plate Ⅰ Partial transverse sections of the twigs of Populus tomentose Carr.showing cambium activity.C.Cambium zone P.Phloem X.Xylem IP.Immature phleom Ⅸ.Immature xylem.1.March 15, cambium entered resuming activity phase, the inclusion in the cambial cell decreased rapidly (thin arrow); × 120;2.March 29, appearance of immature phloem cell, showing enlarging sieve cell (thin arrow); ×120 3.April 21, appearance of immature xylem cell, showing enlarging vessel cell (thin arrow); ×400;4.April 28, increasing of the cell number of cambium and xylem, showing enlarging vessel cell (thin arrow); ×400;5.August 18, showing immature xylem cell (thin arrow)and immature phloem cell (arrow); ×400;6.September 30, disappearance of immature xylem cell and immature phloem cell; ×400;7.December 25, dormant cambium, showing abundant inclusion in the cambial cell(thin arrow); ×400.
崔克明, 利特尔 C H A, 宗德贝格 B. 1992. 欧洲赤松茎部形成层活动和外源IAA对它的影响. 植物学报, 34(7): 515-522. |
崔克明, 罗海龙, 李举怀, 等. 1993. 构树形成层的恢复活动及其过氧化物酶同工酶的变化. 植物学报, 35(8): 580-587. |
崔克明, 魏令波, 李举怀, 等. 1995. 构树形成层的活动周期及其淀粉储量的变化. 植物学报, 37(1): 53-57. |
胡能书, 万贤国. 1985. 同工酶技术及其应用. 长沙: 湖南科学技术出版社, 30~47, 104~110.
|
李正理, 崔克明. 1984. 杜仲再生树皮的不正常发育. 植物学报, 26: 252-257. |
罗立新, 崔克明, 李举怀, 等. 1995. 杜仲形成层活动周期中多糖贮量和淀粉酶同工酶的变化. 北京大学学报(自然科学版), 32: 231-238. |
王雅清, 柴晶晶, 崔克明. 1999. 杜仲次生木质部分化和脱分化过程中酸性磷酸酶的超微细胞化学定位. 植物学报, 41: 1155-1159. DOI:10.3321/j.issn:1672-9072.1999.11.003 |
王雅清, 崔克明. 1998. 杜仲次生木质部导管分子分化中的程序化死亡. 植物学报, 40: 1102-1107. DOI:10.3321/j.issn:1672-9072.1998.12.004 |
薛应龙主编.植物生理学实验手册.上海: 上海科学技术出版社, 1985: 424~432
|
张英伯, 郑槐明, 龙瑞芝, 等. 1982. 八种华北树木形成层季节活动及韧皮部与木质部形成的研究. 林业科学, 18(4): 366-379. |
张仲鸣, 崔克明, 李举怀. 1997. 白皮松形成层的活动周期及其多糖贮量和淀粉酶同工酶的变化. 植物学报, 39(10): 926-932. |
Chaffey N. 1999. Cambium : old challenges—new opportunities. Trees, 13: 138-151. DOI:10.1007/PL00009745 |
Cui K M, Zhang Z M, Li J H. 1995. Changes of peroxidase, esterase isozyme activities and some cell inclusions in regenerated vascular tissues after girding in Broussonetia papyrifera(L. )Vent. Trees, 9: 165-170. DOI:10.1007/BF02418206 |
Evert R F. 1963. The cambium and seasonal development of the phloem in Pyrus malus. Amer. J. Bot., 50: 149-159. DOI:10.1002/j.1537-2197.1963.tb07190.x |
Fahn A.Plant anatomy.3 rd edition.Oxford.England: Pergamon Press, 1982: 291~309
|
Fahn A., Werker E.Seasonal cambial activity.In: Iqbel E.The Vascular Cambium.New York: John Wiley and Sons Ind., 1990: 138~157
|
Libbenga K R, Mennes A M.Hormone binding and its role in hormone action.In: Chadwick C M et al.. Hormones, receptors and Cellular Interactions in Plants.Cambridge: Cambridge University Press, 1986, 194~217
|
Little C H A. 1981. Effect of cambial dormancy state on the transport of[ 1-C14] indol-3-ylacetic acid in Abies balsamea shoots. Can. J. Bot., 59: 342-348. DOI:10.1139/b81-047 |
Luo Lixin, Cui Keming, Li Juhai, et al. 1995. Cambial reactivity and change of peroxidase isozymogram in Eucommia ulmoides Oliv. Chin. J Bot., 7: 150-155. |
Reinecke D M, Bandurski R S.Auxin biosynthesis and metabolism.In: Davies P J.Plant Hormone and Their Role in Plant Growth and Development. Dordrecht: Kluwer Academic Publishers, 1987: 24~42
|
Roberts L W, Aloni P B.Vascular Differentiation and Plant Growth Regulators.Berlin: Spriger-Verlag, 1988: 27, 47, 48 and 78
|
Sachs T. 1981. The control of the patterned differentiation of vascular tissues. Adv. Bot. Res., 9: 152-155. |
Savidge R A, Farrer J L. 1984. Cellular adjustments in the vascular cambium leading to apical grain formation in confers. Can. J Bot., 62: 2872-2879. DOI:10.1139/b84-383 |
Sundberg B, Little C H A, Cui K-M, et al. 1991. Level of endogenous indole-3-acetic acid in the stem of Pinus sylvestris in relation to the seasonal variation of cambial activity. Plant, Cell and Environ., 14: 241-246. DOI:10.1111/j.1365-3040.1991.tb01342.x |
Wodzicki T J, Abe H, Wodzicki A B, et al. 1987. Investigation on the nature of the auxin-wave in the cambial region of pine stems. Plant physiol., 84: 135-143. DOI:10.1104/pp.84.1.135 |