 2002, Vol. 38
2002, Vol. 38文章信息
- 徐有明, 江泽慧, 林汉, 马伟, 羊荣伟.
- Xu Youming, Jiang Zehui, Lin Han, Ma Wei, Yang Rongwei.
- 橡胶树生长轮宽度、木材密度变异及其预测模型的研究
- VARIATION OF GROWTH RING WIDTH AND WOOD BASIC DENSITY OF RUBBERTREE AND THEIR MODELLING EQUATIONS
- 林业科学, 2002, 38(1): 95-102.
- Scientia Silvae Sinicae, 2002, 38(1): 95-102.
-
文章历史
- 收稿日期:2000-10-20
-
作者相关文章


2. 中国林业科学研究院 北京 100091;
3. 海南省大丰农场 琼中 527600
2. Chinese Academy of Forestry Beijing 100091;
3. Dafeng Agricultural Farm of Hainan Province Qiongzhong 527600
橡胶树(Hevea brasiliensis)原产巴西, 因其树皮能分泌乳白色液体生产橡胶, 东南亚地区从19世纪开始大规模种植, 1991年世界上橡胶树人工林面积已达900×104 hm2(Forest Research Institute Malaysis, 1994; 何康等, 1996), 其中90 %在亚洲, 主要为马来西亚、印度尼西亚、泰国和中国等。橡胶树产胶通常在7 ~ 25 a树龄之间, 25 a树龄后胶林需要逐步更新造林。由于橡胶树木材中含有大量的糖类、蛋白质和油类物质, 其木材极易变色发臭腐烂。20世纪80年代以前因无有效的防腐防虫剂和木材加工处理技术, 橡胶木几乎没有商业价值, 多用于薪材或生产木炭(Forest Research Institute of Malaysia, 1994)。实际上橡胶木木材结构细致, 纹理美观, 材色较浅, 呈乳白色或淡黄色, 硬度中等, 加工处理后的橡胶木是生产高档家具、装饰用材、刨花板、中密度纤维板、胶合板等方面的优质材种。目前世界上橡胶人工林每年可生产木材3000×104m3, 其中锯材约800×104 ~ 900×104m3, 这部分木材的有效开发利用不仅大大减少了对热带地区浅色阔叶林的采伐, 而且对保护热带雨林、维护地球生态平衡起到了重要的作用。
我国大规模种植橡胶树开始于20世纪50年代初期(何康等, 1996), 目前海南、云南西双版纳、广东西部和福建南部等地橡胶人工林面积已达61.6×104 hm2, 按照25 a生产周期估算, 每年胶林更新至少可产木材120 ~ 150×104 m3。有关橡胶木木材性质利用研究, 在上世纪80年代中期已有零星报道, 例如蔡则谟(1989)从增加橡胶树品系的抗风折能力方面分别研究了橡胶树有性系(家系)、无性系木材密度的变异规律, 陈传琴等人(1988)报道了胶苗纤维形态、组织比量与茎干冲击的关系; 施振华(1986)对橡胶木防腐防虫试验进行了初步研究, 雷斌(1987)对橡胶木干燥基准进行了初步试验, 但从20世纪90年代中期我国才开始真正重视橡胶木加工利用(刘秀英等, 1997), 橡胶木材性变异与改良、控制机理的研究纳入国家攀登计划有关专题中, 并且我国于1998首次在昆明主办了橡胶木加工利用国际学术研讨会, 胶木兼优品种选育已成为我国橡胶树育种工作者的共识(黄华孙等, 1996)。本文选择我国海南、云南、广东省栽培面积最大、蓄积量最多的2个无性系PR107和PRIM600和“八五”期间大规模推广级的新品系大丰1995(DF95)为试验材料, 研究其株内株间材生长轮宽度、浸提物含量和基本密度变异规律, 建立生长轮宽度和木材基本密度预测模型, 为我国橡胶林分木材材质改良及资源合理开发提供科学依据。
1 材料与方法PR107、PRIM600和DF95 3个无性系15株试材采自海南岛大丰农场橡胶人工林, 地理位置东经109° 44', 北纬19°16', 树龄分别为25 a、25 a和18 a。PR107无性系5株试材胸径分别为29.9 cm、30.0 cm、31.4 cm、32.0 cm和31.8 cm; PRIM600无性系5株试材胸径分别为28.6 cm、29.8 cm、27.3 cm、31.7 cm和35.9 cm; DF95无性系5株试材胸径分别是24.6 cm、25.9 cm、24.7 cm、20.8 cm和20.6 cm。由于橡胶林培育时为降低台风的影响多采用高位截断主干, 其主茎长度多在2.2 ~ 2.5 m之内, 因此每株树木伐倒后于胸高部位截取厚度为2 ~ 3 cm的圆盘3个用于纤维导管形态、化学成份和木材基本密度分析。同时在主干上胸高部位截取两块1 m长左右厚8 cm的中心板用防腐剂简易处理后运回学校, 用于测定木材物理力学性质。
PR107无性系主干高大, 分别从其伐倒木主干基部0.3 m、5.3 m、9.3 m和13.3 m处分别截取圆盘, 研究材性纵向变异规律。同时在阳江农场待更新PR107无性系橡胶林中设置标准地一块, 约1900 m2, 量取胸径后按径阶用生长锥锥取30株木芯, 用于测定、分析林分内株间材性变异规律。
圆盘上由髓心向外量取生长轮宽度后用刨刀逐轮取样, 用排水法测定木材基本密度, 热水浸提物采用造纸GB2677.4 -81中方法进行分析测定。
2 结果与分析 2.1 3个无性系生长轮宽度的变异生长轮宽度反映树木生长快慢, 是林木生长量的重要指标。各个树种生长轮宽度的径向变化既遵守其内在的生长规律, 同时受生长环境的影响。长期以来, 橡胶树一直作为经济作物栽培, 其生长轮宽度变异尚缺乏研究。图 1为橡胶树3个无性系各5株胸高部位生长轮宽度均值的径向变化曲线:由髓心向外生长轮宽度迅速增宽至最大值后, 再向外逐渐减少; 而后随着环境条件改善, 生长轮宽度稍增大后又逐渐变窄, 树皮附近生长轮宽度呈快速下降趋势。这个变化模式与一般用材树种胸径生长规律相同。从图 1来看, 橡胶树3个无性系直径速生期:PR107为4 ~ 9 a, 速生期内生长轮宽度约在0.68 ~ 0.9 cm之间; PRIM600为5 ~ 9 a, 生长轮宽度变化范围为0.68 ~ 0.91 cm; DF95为5 ~ 9 a, 生长轮宽度为0.67 ~ 0.85 cm, 稍低于PR107和PRIM600 2个无性系的生长量。橡胶树开割时间一般在胶林树龄7 a生进行, 即开割期基本上处于橡胶树直径速生期内, 开割2 a后生长轮宽度稍下降, 反映出割胶对橡胶树生长有一定的影响即割胶仅对生长轮宽窄有影响, 对树木直径生长规律无影响, 割胶阶段3个无性系生长轮宽度无显著性差异。橡胶树直径速生期为5 ~ 9 a, 平均生长量约0.68 ~ 0.91 cm, 反映橡胶树生长较快。我国海南、广东、福建橡胶产区多处于台风频发区, 橡胶林多采用高位截干, 主干长度多在2.2 ~ 2.5 m以下, 单株材积小, 单位面积上蓄积量少。如能选用抗风性强的新品系或在无风少风地带不截主干培育胶林, 其更新利用时木材价值将大大增加。事实上随着石油及其产品的开发, 合成橡胶比重逐渐增大, 人们对天然橡胶依赖性已大大降低, 其木材的重要性作为可持续发展的再生资源将会得到人们的广泛重视。建议橡胶产区抓住机遇, 加快培育胶木兼优的新品系, 开发新的经营模式, 培育胶木两用的橡胶林。

|
图 1 橡胶木三个无性系生长轮宽度的变异 Fig. 1 Variation in growth ring width of three clones of rubbertree —◆—PR107, —●—DF95, —▲—PRIM600. |
3个无性系生长轮宽度(y)与树龄(x)间关系可用多项式模型来预测:
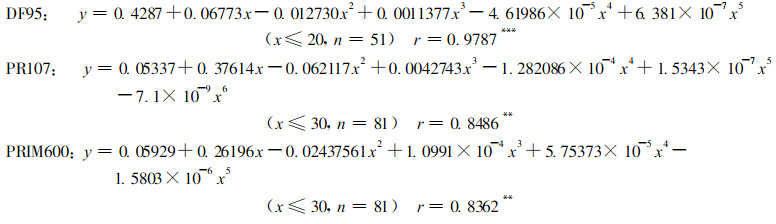
|
经检验分别在0.01或0.001水平上显著, 相对误差可控制在±5 %以内, 这个模型对预估橡胶林分木材蓄积量和确定适宜的营林培育措施具有科学的指导意义。
2.2 浸提物含量的变异木材浸提物是指在细胞腔、细胞腔间隙和细胞壁间的内含物, 能用中性溶剂浸提出来的一共物质总称。尽管浸提物不是木材细胞壁的结构物质, 但其含量高低影响到木材密度大小, 对木材耐腐性、防腐改性处理, 纸浆蒸煮、板材干燥胶合和木材强度等均有一定的影响。通常情况下热带材浸提物含量显著高于温带材。橡胶木作为一种热带材也不例外, 其木材热水浸提物含量较高。图 2为PR107无性系株内不同高度热水浸提物含量的变异曲线, 径向上由髓心向外浸提物含量迅速增加至最大值后稍有下降; 之后又有增大的趋势; 纵向上随着树干高度的增加, 橡胶木热水浸提物含量由基部向上增大, 9.3 m高度截面上浸提物含量最大, 13.3 m高度上稍有降低。0.3 m、1.3 m、5.3 m、9.3 m和13.3 m高度截面上热水浸提物含量变化范围分别为6.65 %~ 9.1 %、6.67 %~ 7.92 %、7.83 %~ 11.72 %、8.65 %~ 10.84 %和8.19 %~ 11.43 %; 平均值依次为7.77 %、7.30 %、8.66 %、9.44 %和9.20 %。变异系数依次为6.75 %、4.34 %、10.37 %、6.29 %和11.53 %。这些数值反映出橡胶树木材热水浸提物含量的确较高, 其株内变异系数较大。方差分析结果显示株内不同高度间浸提物含量差异显著(F =18.58 >F0.05 =2.45)。单因素方差结果显示无性系内株间浸提物含量差异多不显著, PR107无性系内株间浸提物含量F =2.66 < F0.05 =3.14, DF95F =1.85 < F0.05 =3.19, 仅PRIM600无性系F值为6.18 >F0.05 =3.14, 株间差异显著; 无性系间木材浸提物含量差异显著, F值为16.69 >F0.05 =5.14, DF95无性系木材浸提物含量均值为5.87 %, 均小于PR107无性系7.66 %和PRIM600无性系7.8 %, 这对木材密度值大小稍有影响。

|
图 2 橡胶木(PR107)无性系株内木材浸提物含量的径向变异 Fig. 2 Extractives contents variation within trees of PR107 clone -◆-H0.3m, -●-H1.3m -▲-H5.3m, -×-H9.3m, -○-H13.3m |
橡胶林更新采伐后木材极易发生蓝变、黑变、腐烂发臭, 除了热带地区闷热、高湿的环境因素外, 其内在原因与其木材高的浸提物含量及其浸提物中糖、淀粉、蛋白质和油类物质存在有关。因此胶林采伐后木材及时制材、防腐干燥处理是必需的。目前生产上多用水中贮存和端面涂刷油性防腐剂暂时保存木材, 但废水排放和化学防腐剂均对环境有较大的影响, 这点橡胶木产区应引起足够的重视。
2.3 基本密度的变异及预测模型 2.3.1 株内径向变异木材密度是材性的一项重要指标(Pashin, 1980; 成俊卿, 1985), 其变异规律研究对于林木材质改良、选育新品种, 纸浆产量质量评估、实木干燥、改性处理工艺技术的确定及其人造板生产和木材合理利用具有重要的指导意义。图 3为橡胶木PR107无性系5个不同高度木材基本密度的径向变异曲线, 基本密度由髓心向外逐渐增大至最大值后保持相对稳定, 不同高度基本密度稳定开始年限约在5 ~ 8 a左右。树皮附近各个高度木材基本密度均呈不同程度的下降趋势。但树皮附近木材密度值仍高于髓心附近值。蔡则谟、何世强等人(1989)报道橡胶木无性系木材密度的遗传变异时, 其径向变异与本文研究结果稍有差异。差异原因可能在于他们研究中径向取样太少, 仅分为4段, 而找不出完整的变异规律有关。Pashin等人(1980)将阔叶树木材密度径向变异分为3种类型, 本研究橡胶木木材密度径向变异曲线可归于第1类。

|
图 3 橡胶木(PR107)株内木材基本密度的变异 Fig. 3 Variation of basic density within trees of PR107 clone -◆-H0.3m, -■-H1.3m -▲-H5.3m, -○-H9.3m, -*-H13.3m, |
表 1为橡胶树株内3种取样方法木材基本密度的纵向变异的结果。①各个高度从髓心向外在相同的年龄段取样比较, 形成层生理、生长发育阶段相同, 生长轮年龄一致, 反映出不同年度生长环境条件的影响; ②树皮方向由外向内取样, 同一年度形成的木材外部生长环境相同, 但形成层生理、生长发育阶段不同, 反映生长轮年龄的影响; ③各个高度样品取平均值比较。3种方法木材基本密度纵向变异表现出一致的规律, 即随着高度的增加, 木材基本密度增大, 5.3 m高处木材基本密度值最大, 之后又逐渐降低。基本密度的最小值位于树干基部0.3 m处, 这与图 3中不同高度径向曲线一致。表 1中3种不同方法木材基本密度变异系数变优范围为0.3 ~ 8.2 %, 差异不大, 反映出橡胶木株间和株内材质总体上变化小, 橡胶木材质优良。
|
|
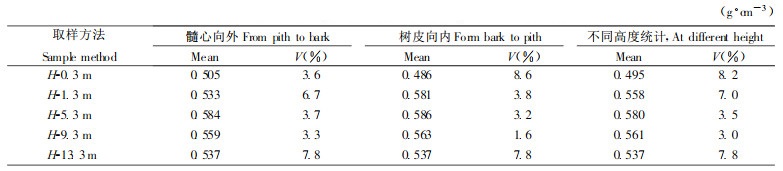
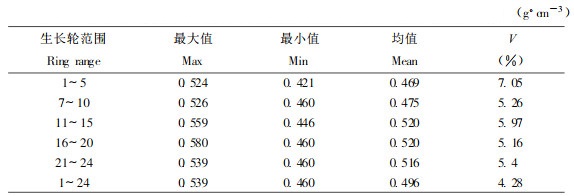
表 2为PR107无性系橡胶林伐前林分内30株样木胸高位置各年轮段木材基本密度的统计分析结果, 木材基本密度均值、最大值和最小值随着生长轮年龄增加表现出一致的变化规律, 即逐渐增大, 达到最大值后又稍微减小, 这与前文3个株内木材基本密度径向变异结果一致。从变异系数大小来看, 林分内株间幼龄期(1 ~ 5轮)基本密度变异较成熟期大, 这与有关理论是一致的。与表 1比较, 橡胶树林分内基本密度变异与株内变异大小说明橡胶树株内木材基本密度变异较小, 材质较为均匀, 有利于提高木材资源加工利用效率, 加工产品质量得到了保证。
|
|
单因素方差分析表明, 3个无性系内株间木材基本密度的差异均达到显著水平, DF95株间F值为11.03 >F0.05 =3.19, 株间基本密度值变化范围为0.529 ~ 0.586 g·cm-3, 变异系数为5.1 %; PR107无性系株间F值为10.94 >F0.05 =3.14, 基本密度变化范围为0.558 ~ 0.605 g·cm-3, 变异系数为4.1 %; PRIM600无性系株间木材基本密度F值为16.46 > F0.05 =3.14, 其基本密度变化范围为0.563 ~ 0.611 g·cm-3, 变异系数为4.2 %。无性系间木材基本密度差异不显著, 无性系DF95木材基本密度均值0.558 g·cm-3, PR107无性系木材基本密度值为0.579 g· cm-3, PRIM600无性系木材基本密度值为0.590 g·cm-3, DF95木材基本密度均稍低于PR107、PRIM600无性系, 可能与DF95浸提物含量值稍低有关。橡胶树无性系内株间基本密度变异大于无性系间的差异, 因此树质改良应着重改良无性系内个体间差异。
2.3.5 浸提物含量对基本密度的影响阔叶树木材的基本密度变异远较针叶木材复杂得多, 其数值大小是导管木纤维、木射线和轴向薄壁细胞等主要解剖分子形态特征值及其组织比量变化的综合反映, 浸提物含量也是影响木材基本密度的重要因子之一(徐有明, 1989)。浸提前DF95无性系3株木材基本密度值分别为0.550 g·cm-3、0.529 g·cm-3和0.586 g·cm-3, PRIM600无性系3株木材基本密度值分别0.596 g·cm-3、0.562 g·cm-3和0.609 g·cm-3, PR107无性系3株胸高盘上基本密度值分别为0.578 g· cm-3、0.599 g·cm-3和0.619 g·cm-3; 浸提后基本密度值:DF95依次是0.515 g·cm-3、0.489 g·cm-3和0.542 g·cm-3, PRZM600依次是0.543 g·cm-3、0.549 g·cm-3和0.516 g·cm-3, 而PR107无性系基本密度值依次为0.539 g·cm-3、0.547 g·cm-3和0.578 g·cm-3。浸提前后木材基本密度值下降幅度:DF95为6.8 % ~ 7.5 %, PRIM600为8.2 %~ 9.8 %, PR107为7.6 %~ 8.7 %, 平均约为8.1 %。这与前文木材浸提物含量变异系数大小基本一致。表 3中线性回归方程说明:橡胶树木材浸提前基本密度(y)和浸提后基本密度值(x)间有很好的拟合性, 决定系数r2变化范围0.8047 ~ 0.9847, 在a =0.01或0.001水平上达到显著水平, 反映出浸提前后木材密度作为育种指标均有很好的代表性, 胶木并用选育抗风性能好的品种或无性系时将木材基本密度作为育种性状是可行的, 可以不考虑浸提物含量的影响。
|
|

木材材性材质预测模型的研究是林木定向培育、材质改良方面重要的发展方向之一。近10 a来, 国际上木材学家已召开3次学术研讨会探讨营林培育与材质模型间关系(Nepveu, 1999)。针叶树、阔叶树株内木材密度径向变异类型各分为3种(Pashin, 1980), 各树种建立同一类型的预测模型几乎是不可能的。目前木材密度预测模型数学方程种类有多项式1)、双曲线(鲍甫成等, 1995)、指数方程(鲍甫成等, 1995)等, 根据图 3橡胶树株内各个高度径向变异曲线, 3个无性系胸高基本密度样本值(y)与生长轮年龄(x)用多项式方程回归叠代计算, 并对方程高次方变量前系数进行微调得到的模型如下:
1) 徐有明.油松木材解剖、物理力学性质的变异及其相互关系的研究。硕士学位论文, 1988。
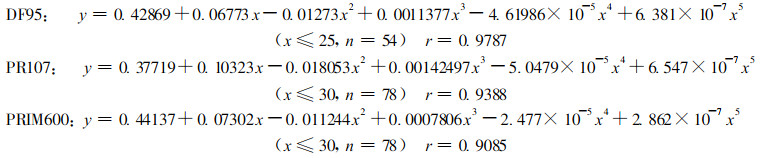
|
经检验上述3个模型在a =0.001水平上检验显著。表 4为橡胶树3个无性系实测值与模型值的比较, DF95多数样本相对误差均小于±3 %, 最大相对误差为4.9 %; PR107样本相对误差小于4 %, 最大相对误差为-7.5 %; PRIM600无性系所有样本相对误差均小于3.5 %, 反映出上述3个模型与实测值误差极小, 3个模型可望在橡胶木材质变异和改良方面发挥重要作用。
|
|

橡胶树3个无性系生长轮宽度径向由髓心向外迅速增大至最大值后递减, 而后随着环境条件变化稍微增宽再趋于减小, 这与一般用材树种胸径生长规律相同。橡胶树直径速生期为5 ~ 9 a, 平均生长量约0.68 ~ 0.91 cm, 反映橡胶树生长较快。我国海南、广东、福建橡胶产区多处于台风频发区, 橡胶林多采用高位截干, 主干长度多在2.2 ~ 2.5 m以下, 单株材积小, 单位面积上蓄积量少。建议橡胶产区抓住机遇, 加快培育胶木兼优的新品系, 选用抗风性强的新品系或在无风少风地带不截主干培育胶林, 开发新的经营模式, 培育胶木两用的橡胶林, 增加其木材的利用价值。
橡胶树木材热水浸提物含量变异有一定规律可循。径向递增到最大值再递减, 而后又递增趋于稳定。纵向上其含量随着高度增加而增大, 再稍微递减。各部位木材热水浸提物含量均较高, 变化范围为6.65 %~ 11.53 %, 远远高于常见阔叶林。橡胶林更新采伐后木材极易发生蓝变、黑变、腐烂发臭, 与其木材高的浸提物含量及其浸提物中糖、淀粉、蛋白质和油类物质存在有关。胶林采伐后, 木材应及时制材、进行干燥与防腐处理。
株内木材基本密度径向变化模式为先递增而后趋于稳定, 树皮附近木材密度有明显的下降。纵向上木材基本密度变化是先递增至最大值, 而后递减。株内木材基本密度较为均匀, 其值范围为0.470 ~ 0.605 g·cm-3, 变异较小, 变异系数范围为3.0 %~ 8.2 %。林分内株间木材基本密度变化范围为0.421 ~ 0.580 g·cm-3, 变化系数在5.16 %~ 7.05 %之间。无性系间木材基本密度差异不显著, 无性系内株间差异显著, 这些说明橡胶树材质改良应重视无性系内个体间差异。橡胶树木浸提前后木材密度值线性相关极显著, 说明浸提前木材密度有很好的代表性, 材质改良研究中可不考虑浸提物的影响。总之橡胶树株间株内木材基本密度变异性不大, 其材质较为均匀。
橡胶树生长轮宽度和木材基本密度与树龄的关系模型为5 ~ 6次多项式方程, 模型在a =0.01或0.001水平上显著, 实测值与模型值误差 < 5 %, 精度高。模型对评估橡胶树木材材积、材质改良研究和木材合理利用具有科学指导意义。
鲍甫成, 江泽慧主编.短周期工业用材林木材性质的研究.世界林业研究, 1995(专集)
|
蔡则谟, 何世强. 1989. 橡胶有性系木材比量的遗传变异. 热带作物学报, 10(2): 31-36. |
陈传琴, 周钟毓. 1988. 一年生胶苗纤维形态和木材组织比重与茎干冲击的关系. 热带作物研究, 3: 13-17. |
成俊卿主编.木材学.北京: 中国林业出版社, 1985
|
黄华孙, 周钟毓. 1996. 我国橡胶树的引种与新品种选育. 热带作物研究, 1: 7-10. |
何康, 黄崇道主编. 1996. 热带北缘橡胶树栽培. 广州: 广东科技出版社, 1-7.
|
雷斌. 1987. 用百度法编制橡胶木材干燥基准初试. 热带作物学报, 8(1): 78-85. |
刘秀英, 蒋明亮. 1997. 中国橡胶木加工利用的调查. 木材工业, 11(5). |
施振华. 1986. 橡胶木防虫防腐试验报告. 林业科学, 22(1): 54-61. |
Forest Research Institute Malaysia. Rubber wood Processing and Utilization. Kepong, Malaysia, 1994: 400
|
Nepveu G. The third workshop"Connection between silviculture and wood quality through modelling approaches and simulation software". December, INPANancy, France. 1999
|
Pashin B. Textbook of Wood technology. 4th Edition, 1980
|