 2002, Vol. 38
2002, Vol. 38文章信息
- 毕晓丽, 洪伟, 吴承祯, 闫淑君, 蓝斌.
- Bi Xiaoli, Hong Wei, Wu Chengzhen, Yan Shujun, Lan Bin.
- 黄山松种群统计分析
- POPULATION STATISTICS ANALYSIS OF PINUS TAIWANENSIS
- 林业科学, 2002, 38(1): 61-67.
- Scientia Silvae Sinicae, 2002, 38(1): 61-67.
-
文章历史
- 收稿日期:2000-11-13
-
作者相关文章


2. 厦门鹭江大学 厦门 361000
2. Xiamen LujiangUniversity Xiamen 361000
黄山松(Pinus taiwanensis)又称台湾松, 产于浙江西天目山海拔700~1200 m、台湾中央山脉海拔750~1800 m、福建中部及西部山区海拔1000~1500 m、安徽省南部、江西、湖南、湖北等省海拔700~1800 m山地, 为我国特有树种, 是产区较高海拔山地重要的绿化和造林树种(《福建森林编辑委员会》, 1993)。种群动态(Population dynamics)是种群生态学的核心, 是研究种群大小或数量在时间、空间上的变化规律。种群统计学(Population statistics)就是对种群的出生、死亡、迁移、性比、年龄结构等进行的统计学研究, 它最初出现在人口统计学上, 现用于一切生物(李博等, 2000)。生命表和存活曲线是研究种群数量动态变化和进行种群统计的重要工具, 生命表结构分析是解释种群变化的前提, 而生命表结构又是这一分析的首要工作, 存活曲线是常见而又直观的方法。通过对种群生命表的编制可得死亡率、损失率等重要参数, 从而可为种群数量统计提供更多的信息(吴承祯等, 2000a)。黄山松种群生态学研究报道较少, 而其生命表分析则鲜见报道(洪伟等, 1997;吴承祯等, 1998)。本研究利用“空间推时间”、“横向导纵向”的方法编制福建省龙栖山自然保护区、屏南县、寿宁县3个不同地方的黄山松种群的静态生命表, 探讨不同环境下黄山松种群的数量动态变化, 同时应用谱分析的方法来分析其种群生长的周期性变化, 旨在为黄山松种群统计学及群落演潜研究提供基础资料。
1 研究区概况及研究方法 1.1 研究区概况龙栖山自然保护区地处福建省西北部的将乐县境内, 东经117°11′~117°21′, 北纬26°23′~26°34′。保护区内山脉走向基本平行于西部的武夷山脉和东邻的戴云山脉, 形成以中、低山为主, 山间盆地零星分布的地形。山体海拔500 m以上, 最高峰1620.4 m。由于地形、植物等因素的影响, 气温、雨量差异较大。海拔1000 m以下地区, 年平均气温在14.6℃~18.8℃, 海拔1000 m以上地区, 年平均气温小于14℃, 年降水量1600~1800 mm。土壤以黄壤为主, 随着海拔的上升, 气温降低, 雨量逐渐增多, 土壤出现红壤—黄红壤—黄壤的规律性变化。植被为原始亚热带常绿阔叶林和常绿针叶林(洪伟等, 2000)。
屏南县位于福建省东北部。地理位置为东经118°41′~119°13′, 北纬26°44′~27°10′, 年平均气温在13℃~18℃, 年总积温4800℃~6500℃, 年降水量在1700~1900 mm范围内, 年均相对湿度78%~83%, 无霜期200~250 d, 为中亚热带山地气候。土壤类型为黄壤或黄红壤, 植被类型为常绿槠类照叶林小区。
寿宁县位于福建省东北部, 东经119°25′, 北纬27°32′。年平均气温14.9℃, 极端最高温35.8℃, 极端最低温-9℃, 年日照时数1792.2 h, 年平均降水量898 mm, 年蒸发量1400 mm, 年相对湿度89%。为中亚热带季风气候, 土壤类型为黄壤或黄红壤, 植被类型为常绿槠类照叶林小区。
1.2 研究方法 1.2.1 调查方法在立地条件、林分状况基本一致的黄山松群落中设立20 m×20 m样方22个(龙栖山区6个, 屏南县为10个, 寿宁县为16个), 每个样方内又设4 m×4 m小样方25个, 共550个小样方, 调查面积为8800 m2。在每个样方内调查株数、量测胸径, 树高和冠幅, 以及每株树在样方中的坐标位置。每一样方挖3个土壤剖面, 分层次取土样带回室内分析。
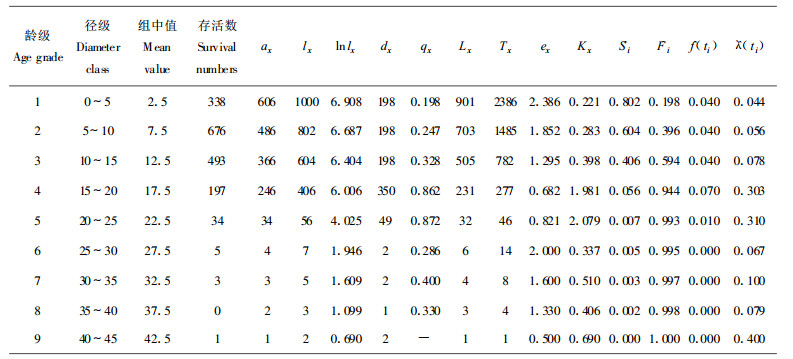
1.2.2 种群年龄结构分析方法树木生长周期长, 不可能追踪所有的个体命运, 因此, 只能通过现实不同年龄阶段的个体数量来推测种群时间上的动态过程, 但由于测定每一种群个体年龄较困难, 故采用空间代时间的方法。即将林木依胸径大小分级, 每级间隔5 cm, 0~5 cm为第1径级, 5~10 cm为第2径级……, 把树木径级从小到大的顺序看作是时间顺序关系, 第1径级对应I龄级, 第2径级对应第Ⅱ龄级, 如此一一对应, 统计各龄级株数, 编制黄山松种群静态生命表, 进而分析其动态变化。
1.2.3 生命表编制特定时间生命表一般包含如下栏目:x为单位时间年龄等级的中值; ax为在x龄级内现有个体数:lx为在x龄级开始时标准化存活个体数(一般转化为1000);dx为从x到x+1龄级间隔期内标准化死亡数; qx从x到x+1龄级间隔期间死亡率; Lx为从x到x+1龄级间隔期间还存活的个体数; Tx为从x龄级到超过x龄级的个体总数; ex为进入x龄级个体的生命期望寿命; Kx为消失率(损失度) (祝宁等, 1994)。
生命表中各项都是相互关联的, 可以通过实测值ax或dx求得, 其关系如下:
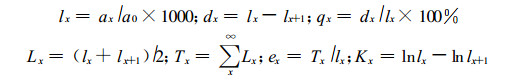
|
同时本文引入生存分析中的4个函数项目于黄山松种群生命表栏目中, 即生存函数S (t)、积累死亡率函数F (t)、死亡密度函数f (t)、危险率函数λ (t) (冯士雍, 1982;1982;1983)。

|
波动出现于所有的植被中, Veblen等通过老龄林结构和动态分析认为, 优势种的林冠更替是周期循环的, 而不是一个连续发展过程。天然更新过程是不同林分或同一林分内不同年龄林木的更替过程。谱分析则是探讨这种分布的波动性和年龄更替过程的周期性的数学工具(伍业钢等, 1988)。黄山松种群天然更新过程的动态是通过黄山松不同龄级的株数分布波动表现的。因此, 本文对黄山松种群进行数量动态的谱分析, 以探讨谱分析方法在种群统计学中的应用。
谱分析是Fourier级数的展开。Fourier证明过复杂的周期现象可以由不同振幅和相应的谐波组成, 写成正弦波形式:
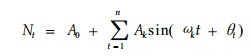
|
式中, A0为周期变化的平均; Ak (k=1, 2, 3, …, p)为各谐波的振幅, 标志其所起的作用大小, 其值的差异, 反映了各周期作用的大小的差别; ωk及θk分别为谐波频率及相角; Nt为t时刻种群大小(吴明作等, 2000)。
将种群各年龄个体分布视为一个时间系列t, 以Xt表示t年龄序列时个体数; n为系列总长度; p=n/2为谐波的总个体数为已知; T为正弦波的基本周期即时间系列t的最长周期, 即资料的总长度, 这里T=n是已知的。则可以利用下式来估计Fourier分解中的各个参数, 即:
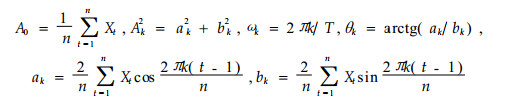
|
由于静态生命表用同一时期收集到的种群中所有个体的径阶编制而成, 它反映了多个世代重叠的年龄动态历程中的一个特定时间, 而不是对同生群的全部生活史追踪(Silvertown, 1987;苏智先等, 1993;刘金福, 1999;吴承祯等, 2000b), 因此, 在生命表的编制中有可能会出现死亡率为负的情况, 对此, 本文采取文献(江洪, 1992)中的匀滑技术进行处理, 经匀滑修正后得ax (表 1~3), 然后, 再根据此编制出黄山松种群特定时间生命表。
|
|

|
|
|
|
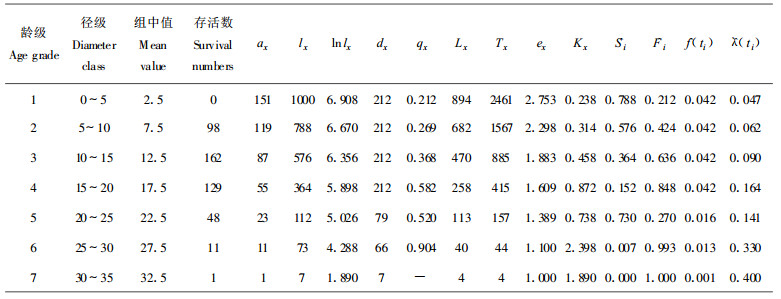
以匀滑技术处理后得到的ax值, 根据静态生命表的编制方法, 同时引入生存分析中的4个生存函数来编制不同地区黄山松种群静态生命表。由表可知3个地区的黄山松种群个体的存活数都随着年龄的增加降低, 但其死亡的强度或速率并不一致(表 1~3)。相应的黄山松的林木生长时间长短不一, 其中为龙栖山黄山松种群最长为11个龄级, 屏南县黄山松种群次之为9个龄级, 寿宁县黄山松种群最短为7个龄级。这与黄山松所处的环境、气候条件等因素有关, 龙栖山位于闽西北地区, 气候较适宜, 利于黄山松生长; 而屏南和寿宁2县均位于闽东北地区, 地处沿海县海拔较高, 易受台风影响, 这些因素使得该区黄山松与龙栖山黄山松在生长、生命过程中存在一定的差异。
2.2 黄山松种群的存活曲线本文以存活量lx为纵坐标, 以径级相对的年龄级作横坐标, 作存活曲线图(图 1)。Peal (1923)在对死亡率进行分类时, 首先提出了种群存活曲线。Deevey (1947)随后把存活曲线分成3型:Ⅰ型(又称a型)存活曲线呈凸型, 表示种群的大多数个体均能实现其平均的生理寿命, 在达到平均寿命时, 几乎同时死亡; Ⅱ型(又称b型)存活曲线呈对角线型, 表示各龄级具有相同的死亡率; Ⅲ型(又称c型)存活曲线呈凹型, 表示幼体死亡率高, 以后的死亡率低而稳定。据Deevy的划分, 由图得知, 本研究中的黄山松种群的存活曲线应属于Ⅱ和Ⅲ型之间的过渡类型。从图可以看出3个不同地区的黄山松种群的存活曲线均为同一种类型, 反应了黄山松种群整体的数量动态变化趋势及结构特征, 其中黄山松幼年的存活值相对较高, 死亡数较少, 而从4~7龄级阶段存活率开始下降, 最终达平衡稳定状态。这是由黄山松的生物学特性决定的, 黄山松耐寒冷, 抗风力强, 耐瘠薄土壤, 幼苗生长健壮。

|
图 1 不同地区黄山松种群的存活曲线 Fig. 1 Survival curve of Pinus taiwanensis population in different areas —◆ —龙栖山Longxi Mountain, —■—屏南县Pingnan, —●—寿宁县Shouning. |
以死亡率为纵坐标, 以径级相对的龄级为横坐标, 作死亡率曲线(图 2)。死亡率曲线反映了不同地区黄山松种群死亡率的动态变化, 从整体上讲3个地区的黄山松种群都存在两个死亡率高峰: (1)在近成熟前期, (2)成年阶段。幼年死亡率低是因为气候条件、地理环境的适宜及黄山松种群的生物学特性所决定的, 其幼苗耐性强、生长好。而到近成熟时期, 黄山松长势良好, 对阳光、水分等资源的需求加大, 种内竞争增大, 出现第1个死亡高峰。到了成年阶段, 又出现1次小的死亡高峰, 这可能是由于种间的竞争的存在。从图 2中还可以看到, 不同地区的死亡率曲线存在明显的差异:屏南、寿宁2县的黄山松死亡率高峰出现较早, 而龙栖山区的黄山松死亡率高峰则要滞后1~2个龄级; 在第1龄级时的死亡率的大小是寿宁县 > 屏南县 > 龙栖山区; 死亡率最大值为寿宁县(0.904) > 屏南县(0.872) > 龙栖山区(0.743);寿宁县的黄山松种群最早结束整个生活史, 龙栖山地区的黄山松种群生长情况最好。其原因可能是与3个地区所处的地理环境和气候条件有关, 寿宁县、屏南县位于气候独特自成一区的鹫峰山地区, 因平均地势高, 年平均气温比全省年平均气温低3℃~4℃, 极端低气温最低。黄山松性喜高海拔温凉、湿润气候, 因此, 在龙栖山区黄山松种群自然生长条件最好, 此处的生长龄级为最长。另一方面黄山松种群在成年阶段的死亡率相对较低, 则反映了黄山松种群的成年阶段对环境的敏感性大于生物竞争, 即对生物竞争的耐性大, 这与祝宁等(1994)对刺五加种群的研究结果是一致的。

|
图 2 不同地区黄山松种群的死亡率曲线 Fig. 2 Mortality rate curve of Pinus taiwanensis population in different areas — ◆ —龙栖山Longxi Mountain, —■—屏南县Pingnan, —●—寿宁县Shouning. |
同样以“空间代时间”的方法将不同地区的黄山松以径级代龄级, 得数据长度n即为所分径级数, 以n×5代以基波的基本周期年限。因所用资料各龄级个体数量相差甚大, 在计算时进行了对数化处理, 即以X′t=ln (Xt+1)代换公式中的Xt, 利用谱分析中的公式计算各种情况下各个波形的振幅Ak (k=1, 2, 3, …p; p=n/2), 结果见表 4, 表 4中n为数据总长度; A1为基波; A2~A7为各个谐波。每个谐波的周期分别是基本周期(相对于径级)的1/2, 1/3, …1/p, AK值的大小差异反映了各周期作用大小的差别。
|
|
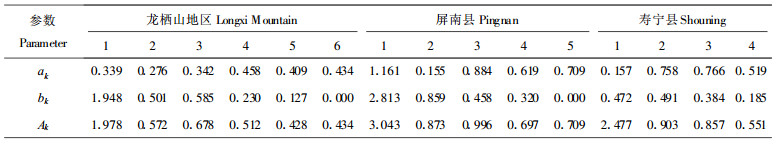
从表 1~3中可以看出, 所调查的3个地区的黄山松种群年龄较小, 种群属于典型的增长型种群的特征, 基波即是表现了基本周期的波动, 其周期长度为种群本身所固有, 由种群波动特性决定。由表 4可知, 各调查地区的AK值均以A1最大, 表明黄山松种群的数量动态变化过程受基波影响很明显。与天然更新过程的周期波动一样, 黄山松生长动态也表现出大周期内的小周期迭加, 如表 4中的A3说明此时为一个小周期, 这是黄山松高生长达到林层高度时, 林木产生分化现象时的数量变动; 林分由郁闭变为稀疏, 数量进行调节, 有利于种群的发展, 使其能自我维持稳定。不同地区的黄山松种群龄级周期长短不等, 但整体上表现出大周期内有小周期的波动, 这种波动使得种群得以维持高的优势度, 这与黄山松纯林即达到群落演替的顶极是一致的。
2.5 亏损度曲线由表 1计算得的值Kx为纵坐标, 以径级相对的龄级为横坐标作Kx曲线(图 3), 并计算出3个地区中黄山松在7龄级时的总亏损度(K total)。结果为寿宁(6.908) > 屏南(5.809) > 龙栖山(3.556), 这与3地区的黄山松种群的生长周期不同有相似之处, 寿宁县的黄山松的生长周期最短, 所以在7龄时大多已死亡, 总亏损度最大。

|
图 3 不同地区的黄山松种群的KX值曲线 Fig. 3 Value curve of Pinus taiwanensis population in different areas —◆ —龙栖山Longxi Mountain, —■—屏南县Pingnan, —●—寿宁县Shouning. |
生存分析中的4个函数的估计值说明了黄山松种群具有前期增长, 后期稳定的特点, 树木死亡随年龄的增加而增加的特点, 仅以3地区的危险率函数值作图(图 4)为例, 同死亡率曲线相比, 结果是一致的。

|
图 4 不同地区黄山松种群的危险率函数曲线 Fig. 4 Hazard rate function curve of Pinus taiwanensis population in different areas —◆ —龙栖山Longxi Mountain, —■—屏南县Pingnan, —●—寿宁县Shouning. |
由黄山松种群生命表分析表明, 其种群的存活曲线为DeeveyⅡ-Ⅲ型曲线, 3个地区的曲线变化趋势是一致的, 从整体上反映了黄山松种群数量动态变化趋势, 这是由黄山松的生物学特性及抗病、虫能力强所决定的。黄山松纯林时达到群落演替的顶极, 相对稳定, 生长后期死亡率高, 这时才有利于阔叶树种的入侵, 使群落由针叶林向针阔混交, 进而进入阔叶林的演替过程。
不同地区的黄山松种群都有两个死亡率高峰出现, 高峰期出现的时间格局不同, 死亡的强度也不同, 以出现的早晚顺序、死亡强度为寿宁县 > 屏南县 > 龙栖山区, 这是不同地区的地理位置、物理环境、气候等多种因素综合作用的结果。亏损度的情况也是如此。
新引入生命表栏目中的几个函数很好地说明种群的结构和动态变化。生存函数与累积死亡率函数、死亡密度函数及危险率函数之间是相互换算的, 因此在实际中可按具体情况选择使用。生存函数曲线同生存曲线相比, 是任意时间t (龄级)的函数, 曲线同实际情况较之生存曲线更直观、具体。对最能体现本省黄山松种群生长情况的屏南县而言, 其危险率曲线、死亡率曲线同损失度曲线变化趋势相似, 都是前期变化幅度大, 后期变化幅度小。同时, 根据黄山松种群特定的生存数据, 可对生存函数(或其他相关函数)进行估计和检验, 说明生存函数在黄山松种群生命表分析中有着很高的实际应用价值。
波动出现于所有植被中, 谱分析方法适用于增长型种群, 其结果表明黄山松种群的周期长度为龙栖山地区 > 屏南县 > 寿宁县, 同时大周期内有小周期的波动。优势种的林冠更替是周期循环的, 而不是一个连续发展的过程, 其波动使其自我维持稳定性, 成为生存区域内的稳定森林群落, 谱分析可用于表明这种机制。
冯士雍. 1982. 生存分析Ⅰ. 数学的实践与认识, (3): 72-80. |
冯士雍. 1982. 生存分析Ⅱ. 数学的实践与认识, (4): 64-74. |
冯士雍. 1983. 生存分析Ⅲ. 数学的实践与认识, (1): 70-76. |
《福建森林》编辑委员会编著.福建森林.北京: 中国林业出版社, 1993: 43~47
|
洪伟, 吴承祯, 林成来, 等. 1997. 龙栖山黄山松种群优势度增长规律研究. 福建林学院学报, 17(2): 1-5. |
洪伟, 吴承祯, 林成来, 等. 2000. 福建龙栖山森林群落林窗边缘效应研究. 林业科学, 36(2): 33-38. |
江洪. 1992. 云杉种群生态学. 北京: 中国林业出版社, 7-139.
|
李博主编.生态学.北京: 高等教育出版社, 2000, 46
|
刘金福, 洪伟. 1999. 格氏栲种群增长动态预测研究. 应用与环境生物学报, 5(3): 247-253. DOI:10.3321/j.issn:1006-687X.1999.03.003 |
Silvertown J W.祝宁, 王义弘, 陈文斌译.植物种群生态学引论.哈尔滨: 东北林业大学出版社, 1987, 19~74
|
苏智先, 王仁卿主编.生态学概论.北京: 高等教育出版社, 1993, 41~44
|
吴承祯, 洪伟, 林成来. 1998. 黄山松种群数量动态研究. 浙江林学院学报, 15(3): 274-279. |
吴承祯, 洪伟, 谢金寿, 等. 2000a. 珍稀濒危植物长苞铁杉种群生命表分析. 应用生态学报, 11(3): 333-336. |
吴承祯, 洪伟著.杉木数量经营学引论.北京: 中国林业出版社, 2000b, 125~130
|
吴明作, 刘玉萃. 2000. 栓皮栎种群数量动态的谱分析与稳定性. 生态学杂志, 19(4): 23-26. DOI:10.3321/j.issn:1000-4890.2000.04.005 |
伍业钢, 韩进轩. 1988. 阔叶红松林红松种群动态的谱分析. 生态学杂志, 7(1): 19-23. |
祝宁, 臧润国. 1994. 刺五加种群生态学的研究Ⅱ.刺五加的种群统计. 应用生态学报, 5(2): 237-240. |