 2002, Vol. 38
2002, Vol. 38文章信息
- 杨小波.
- Yang Xiaobo.
- 南亚热带4个不同演替阶段树种苗木环境适应性研究
- STUDY ON ADAPTABILITY TO ENVIRONMENT FOR 4 TREE SPECIES AT DIFFERENT SUCCESSIONAL STAGES GROWN IN SOUTH SUBTROPICAL ZONE
- 林业科学, 2002, 38(1): 56-60.
- Scientia Silvae Sinicae, 2002, 38(1): 56-60.
-
文章历史
- 收稿日期:1999-10-21
-
作者相关文章


光和水一直被认为是植物种间相互替代, 尤其是次生林在发展成气候顶极群落的演替过程中, 植物相互替代的主要因素(Bazzaz, 1979)。一般来说, 形成树荫后, 林下的苗木都是生活在一种光强度不足的逆境之中, 某一物种是否能在群落中生存、生长和发育取决于其苗木是否能适应这种弱光的环境。为了定量研究这种适应性, 国外较多学者常选择不同森林演替阶段的优势种(单优种或多优种)苗木, 利用不同的生长和生理指标从生态位宽度和生态位重叠等方面开展研究(Bazzaz, 1987;Parrish, 1982)。目前结合植物生理和生态学的手段开展不同森林次生演替阶段的优势种苗木的生态位研究已经是揭示森林次生演替机理的重要研究内容之一(钟章成, 1995)。
过去广大学者的几十年群落演替的定性和定量的研究, 认为我国亚热带广大地区的森林次生演替规律是:马尾松为优势的常绿针叶林→马尾松为优势的常绿针阔叶混交林→阔叶树为优势的常绿针阔叶混交林→常绿阔叶林。同时广大学者也对不同地区、不同演替阶段的植物群落优势种群动态也开展了深入的研究(董鸣等, 1986;刘雄思等, 1987;王伯荪, 1982)。但是结合生态位理论, 从植物生理生态学角度对这个次生森林演替序列开展优势种苗木对环境的适应性研究的报导较少, 本研究是在群落调查的基础上开展这方面研究的。
1 研究材料与方法 1.1 研究材料研究材料为马尾松(Pinus massoniana), 演替先锋树种; 荷木(Schima superba)、藜蒴(Castanopsis fissa), 演替过渡种; 黄果厚壳桂(Cryptocarya concinna), 演替后期种的苗木。1 a生苗木来自广东黑石顶自然保护区。
1.2 研究方法 1.2.1 培养方法分别在校园里分全日照、每天直照光1 h (人工挡板控制, 10:00~11:00 Am为太阳直照时间, 类似林窗环境)和室内窗边等3组光环境中土培上述4种苗木, 培养1 a后, 对室内窗边土培的苗木又分成3组, 其中一组隔3 d供水, 一组隔6 d供水, 一组隔9 d供水(共27 d); 在室内窗边光环境中, 用不同浓度梯度的矿物质培养液培养同样的苗木, 时间为3周, 培养液的浓度为基本培养液(华东师范大学生物系, 1980)的100%、75%、50%、25%、15%和10%, pH均为5.6, 共6组; 用基本培养液调制成不同pH值梯度(pH为7.6、6.8、5.6、5.1和4.8)的培养液培养同样的苗木, 时间为3周, 共5组。
1.2.2 测定指标和方法用LI-6200便携式光合作用仪测定各苗木的光合作用强度, 同时测定室内窗边不同时间供水的3组苗木的土壤含水量; 对全日照、每天直照光1 h和室内窗边等光环境中土培的4种苗木, 用14C示踪法(浓度为100 μci的14C-葡萄糖液, 每处理叶为300 μL, 处理后3 d, 分处理叶、顶芽、处理叶的下一轮叶(简称下叶)、茎和根取样, 烘干恒重, 取20 mg, 测定其脉冲数(cmp·mg-1))测定光合产物的输出量。以所测定的光合强度值为指标, 用生态位理论分析这些苗木对光、矿物质浓度和培养液pH的适应程度(余世孝, 1993)。
2 研究结果与分析 2.1 苗木对不同的环境条件的生态位宽度与重叠分析以物种苗木在不同的环境中的净光合作用速率为指标, 用物种生态位的宽度和重叠理论来分析不同苗木对环境条件的适应性。表 1是这些苗木的生态位宽度和重叠度测定值。从表 1可以发现, 演替过渡种藜蒴对pH值的变化适应性较强, 生态位宽度为0.96, 其它3个物种稍弱一些。4个物种的生态位宽度大小顺序是藜蒴 > 马尾松 > 黄果厚壳桂 > 荷木, 在演替序列上没有一定的规律; 演替过渡种藜蒴和荷木对矿物质浓度梯度变化适应性较强, 生态位宽度均为0.86;先锋树种马尾松和演替后期种黄果厚壳桂对矿物质浓度梯度变化适应性比较弱, 分别为0.78和0.76。从而反映了森林演替过渡种比先锋树种和演替后期种有较强的对矿物质营养浓度等条件变化适应性。但是对木壤水资源轴来说, 4个物种苗木的生态位宽度差别较大, 大小顺序是马尾松 > 荷木 > 藜蒴 > 黄果厚壳桂, 马尾松明显比演替过渡树种有较强的适应水环境变化的能力, 过渡树种又明显比演替后期树种有较强的适应水环境变化的能力, 有明显的演替序列规律性。4个树种苗木的生态位重叠度都较高, 说明在森林演替过程中, 各阶段的树种对资源的竞争较激烈。
|
|

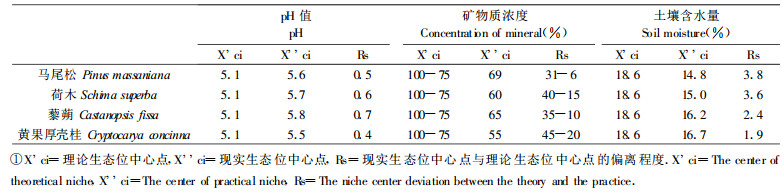
一物种的理论生态位中心定义为在生态位空间具有最佳的适应位置, 因此物种在某一资源轴上的某一生理指标占有最佳比例的相应资源状态为物种在这一资源轴上, 这一生理反应的理论生态位中心点。光合作用强度的测定结果表明, pH=5.1、矿物质浓度为75%~100%和土壤含水量为18.6%分别是这3个资源轴上光合作用强度这一指标占有最佳比例的相应资源状态(表 2), 因此可以认为pH 5.1、矿物质浓度为75%~100%和土壤含水量为18.6%分别是这3个资源轴上理论生态位中心点。表 3为不同苗木的现实生态位中心点与理论生态位中心点的偏离程度测定结果。从表 3可以看出, 对pH值来说, 黄果厚壳桂的偏离值最小, 较适合在pH 5.1左右的环境中生存和生长, 其它的几个物种是马尾松 < 荷木 < 藜蒴; 对土壤含水量来说, 同样是黄果厚壳桂的偏离值最小, 该苗木只能生存和生长在土壤含水量较高的中生环境中; 对土壤含水量的变化(变干旱)的适应能力最强的是马尾松, 其次是荷木、藜蒴, 黄果厚壳桂最小。但是对矿物质营养来说, 这几个物种苗木对它的变化的适应能力顺序是黄果厚壳桂 > 荷木 > 藜蒴 > 马尾松。当然这仅是光合作用这一指标的反应情况。
|
|

|
|
从表 4可以看到, 演替先锋树种马尾松的苗木是在全日照的光环境中有利于光合产物的输出, 而且运往生长顶芽的比例较大, 占总输出量的17.0%, 属正常的生理活动; 但是在弱光或“林窗”的环境中, 光合产物的输出比例较小, 运往生长顶芽的比例更小, 仅分别占总输出量的7.5%和5.8%, 不利于光合产物的输出; 演替过渡种荷木和演替后期种黄果厚壳桂都是在有一定的遮光的环境中, 特别是在“林窗”的环境中, 较有利于光合产物的输出, 强光不利于光合产物的输出。藜蒴的情况比较特殊, 在全日照和“林窗”的环境中, 光合产物的输出都比较小, 而在室内窗边的弱光环境中的输出量稍大一些。总的来说, 弱光对森林演替先锋树种马尾松的光合产物的输出与分配是不利的, 而强光却不利于森林演替过渡种和演替后期种苗木的光合产物的输出与分配。
|
|

本文用净光合作用速率为指标从生态位宽度、重叠和理论生态位中心与现实生态位中心的偏离程度等方面来探讨不同演替阶段森林群落的优势树种苗木对环境适应性的问题, 并采用同位素技术研究了光环境与苗木光合产物的输出和分配的关系。研究的结果表明, 森林演替先锋树种马尾松的苗木对土壤含水量变化(变旱)和全日照的强光有较强的适应性, 易于占领土壤含水量变化(变旱)较强烈、光照强的荒坡荒地; 演替后期种黄果厚壳桂对土壤含水量的变化的适应性较差, 生态位宽度和偏离值都较小, 该苗木较适宜在土壤含水量为18.4%左右的中生环境中生长; 仅从生态位宽度大小顺序:藜蒴 > 马尾松 > 黄果厚壳桂 > 荷木, 看不出各物种对pH值的变化有规律性的适应, 但从偏离值的大小顺序就可以看出演替过渡种藜蒴对pH值的变化适应性较强, 演替后期种黄果厚壳桂的适应性较差, 该苗木较适宜在pH5.1左右的中性环境中生长; 从生态位宽度与生态位中心偏离值都表现出演替过渡种藜蒴和荷木对矿物质浓度梯度变化适应性较强, 这与它们苗木有较发达的根系相关密切, 从而反映了森林演替过渡种比先锋树种和演替后期种有较强的对矿物质营养浓度等条件变化适应性。同时反映了生态位宽度与生态位中心偏离值相结合研究这方面的问题比仅从生态位宽度来得理想。4个树种苗木的生态位重叠度都较高, 说明在森林演替过程中, 各阶段的树种对资源的竞争是比较激烈的。当然, 由于生态位中心偏离值理论是刚发展起来的(Parrish, 1982), 因此尚未有人用生理指标计算物种在不同资源轴上的理论生态位中心与现实生态位中心及其偏离值, 本文在这方面仅做了一些尝试性工作, 有待于进一步发展。同时光合产物输出与分配规律的研究还表明了, 弱光对森林演替先锋树种马尾松的光合产物的输出与分配是不利的, 而强光却不利于森林演替过渡种和演替后期种苗木的光合产物的输出与分配。
董鸣. 1986. 缙云山马尾松种群数量动态研究. 植物生态学与地植物学学报, 10(4): 283-293. |
刘雄思, 王伯荪. 1987. 黑石顶自然保护区植被分类系统和主要类型及分布. 生态科学, (合刊): 19-34. |
王伯荪, 马曼杰. 1982. 鼎湖山自然保护区森林群落的演变. 热带亚热带森林生态系统研究, 1: 142-158. |
余世孝. 1993. 基础与现实生态位及其中心点的涵义与测度. 中山大学学报(自然), 32(4): 69-80. |
钟章成. 1995. 四川大头茶若干生态问题的研究. 植物生态学报, 19(2): 103-114. DOI:10.3321/j.issn:1005-264X.1995.02.005 |
Bazzaz F A. 1979. The physiology of ecology of plant succession. Ann Rev. Ecol. Syst, 10: 351-371. DOI:10.1146/annurev.es.10.110179.002031 |
Bazzaz F A. Experimental studies on the evolution of niche in successional population. In colonization in plant population ecology, ed A. J Gray, . Blackwell scientific publication Oxford London : 1987 : 245~271
|
Parrish J A D. 1982. Responses of plants from three successional communities to a nutrient gradient. J. Ecol, 70: 233-248. DOI:10.2307/2259876 |