 2002, Vol. 38
2002, Vol. 38文章信息
- 喻理飞, 朱守谦, 叶镜中, 魏鲁明, 陈正仁.
- Yu Lifei, Zhu Shouqian, Ye Jingzhong, Wei Luming, Chen Zhengren.
- 退化喀斯特森林自然恢复过程中群落动态研究
- DYNAMICS OF A DEGRADED KARST FOREST IN THE PROCESS OF NATURAL RESTORATION
- 林业科学, 2002, 38(1): 1-7.
- Scientia Silvae Sinicae, 2002, 38(1): 1-7.
-
文章历史
- 收稿日期:2001-02-21
-
作者相关文章


2. 南京林业大学 南京 210037;
3. 茂兰国家级自然保护区管理局 荔波 558400
2. Nanjing Forestry University Nanjing 210037;
3. The Management of Maolan National Reserve Libo 558400
南方喀斯特森林是一种脆弱的生态系统, 因人类不合理开发利用, 已大面积退化, 甚至形成“石漠化”, 严重威胁农业生产环境乃至人类生存, 退化喀斯特森林恢复与重建意义重大。但由于喀斯特环境特殊性, 恢复与重建困难, 成为我国南方退化生态系统恢复与重建研究的难点与重点。
相对于对地带性植被如常绿阔叶林的研究而言, 对喀斯特森林的研究甚少, 最早可追溯到上世纪40年代(郭魁士, 1940;侯学煜, 1946;1952), 80年代对喀斯特森林顶极群落特点、属性的讨论成为焦点(王献溥, 1981;胡舜士, 1982;周政贤, 1987;方任吉, 1988;肖育檀, 1988;屠玉麟, 1989;吴春林, 1991), 进入90年代, 一方面围绕顶极群落结构功能、主要种群动态深入研究(朱守谦, 1993;1995;1997;杨汉奎, 1991;梁士楚, 1992;杜道林, 1996;喻理飞, 1998a), 另一方面, 开展了对喀斯特森林退化原因、过程、生境条件及树种适应性研究(杨继镐, 1990;徐樵利, 1993;何道泉, 1993;张祝平, 1993), 并提出退化喀斯特森林自然恢复途径和恢复评价(苏宗明, 1990;喻理飞, 1998b;2000)。但对退化喀斯特森林自然恢复的生态学过程缺乏系统研究, 因此, 研究恢复过程中群落动态变化, 加深对其自然恢复生态学过程的理解, 对恢复与重建喀斯特森林生态系统具有重要的理论意义和现实意义。
1 研究区域概况本研究在贵州茂兰国家级自然保护区内进行。地理位置为东经107°52′~108°05′, 北纬25°09′~25°20′。年均温18.3℃, ≥10℃积温5767.9℃; 全年降水量1320.5 mm, 集中分布于4~10月; 年均相对湿度80%;年均霜日7.3 d; 全年日照时数1272.8 h, 日照百分率29%。属中亚热带季风湿润气候, 有利于林木生长。区内主要出露岩石为纯质石灰岩和白云岩, 属裸露型喀斯特地貌, 与常态地貌相比, 生境复杂多样, 有石面、石沟、石洞、石槽、石缝、土面等多种小生境类型, 其生态因子变化很大(朱守谦, 1993)。土壤以黑色石灰土为主, 土层浅薄且不连续, 剖面构型多为AF-D型、A-D型。地表水缺乏, 土体持水量低, 土壤富钙和富盐基化, pH6.5~8.0, 有机质含量高。
保护区多数地段是中亚热带原生性喀斯特森林, 其顶极群落为常绿落叶阔叶混交林(周政贤, 1987), 也有不同退化程度的演替群落, 对退化群落自然恢复的研究有很强的代表性。
2 研究方法 2.1 调查方法本研究采用样地法取样。在各演替阶段设置样地, 取样面积分别为:顶极群落样地30 m×30 m, 乔木样地20 m×30 m, 灌木样地16 m×10 m, 草本群落4 m×5 m。每个顶极群落样地分为10个6 m×15 m乔木样方, 在每个乔木样方内设置1个4 m×4 m灌木样方和1个1 m×1 m草本样方。每乔木样地分为10个6 m×10 m乔木样方, 在每个乔木样方内设置1个4 m×4 m灌木样方和1个1 m×1 m草本样方。每个灌木样地分为10个4 m×4 m灌木样方10个1 m×1 m草本样方。调查记录每个样方内乔、灌木及幼树的种类、数量、胸径、地径、高度、冠幅、盖度, 并判定每株树木是实生或萌生起源; 草本植物调查其种类、多度、平均高、盖度。共获顶极群落样地、乔木样地、灌木样地、草本群落样地分别为6、15、25、8个。
群落生物量按乔木层、灌木层、草本层分别调查。前二者采用样本法, 根据调查结果, 按树种、层次、胸径、地径选取样木, 用收获法测定样木树干(含树皮)、枝、叶的鲜重。乔木层、灌木层分别获取样本30、119株。在每块样地内设置3个1 m×1 m草本层样方, 采用刈割法获取草本层地上部分鲜重。取样品在80℃恒温烘至恒重, 计算含水率并换算成干重。
2.2 分析方法本研究采用以“空间代替时间” (D.Mueller-Dombois et al., 1986)的方法, 将54个群落样地通过样地聚集、聚集团联结、样地定位等一系列步骤, 构建退化群落自然恢复的演替系列, 并用最优分割法将退化群落自然恢复过程分为草本群落阶段、草灌群落阶段、灌丛灌木阶段、灌乔过渡阶段、常绿落叶阔叶林阶段和顶极群落阶段6个演替阶段(喻理飞, 1998b)。
喀斯特地区生境复杂, 能适应同一演替阶段生境的物种不是一个而是具有相同适应性的多个物种构成的种组。根据退化群落在自然恢复过程中主要组成物种优势地位的变化, 划分适应等级种组, 具体方法如下:分别各演替阶段选择一典型样地, 以种群为实体, 种群重要值和树高为属性, 建立种群在群落恢复过程中优势地位矩阵, 采用对应分析法(COA) (Jhon, 1990)进行分析, 根据种群空间位置分布状况, 划分为先锋种、次先锋种、过渡种、次顶极种和顶极种5个适应等级种组; 再根据54个样地中所有种群与各种组中树种的联结性和协变关系, 分别归为不同种组1)。
1) 喻理飞.退化喀斯特森林自然恢复的生态学过程研究.南京林业大学博士学位论文, 1998.
相对生长关系被广泛运用于群落生物量的估测。采用W=a (D2H) b的回归式分别建立林木地上部分生物量(W)与树高(H)、胸径(D胸径)或地径(D地径)的关系式为:
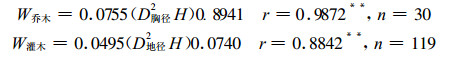
|
群落中乔木层、灌木层生物量分别按上述回归式及其相应的株数求得, 草本层生物量采用刈割法实测获得。
物种多样性采用以下指标度量:Shannon-Wiener多样性指数(H′), H=3.3219[lgN- (1/N) (∑nilgni) ]; 生态优势度用Simpon指数(C), C=∑ni (ni-1) /N (N-1);均匀度采用以Shannon-Wiener多样性指数为基础的均匀度指标(J), J=[lgN- (1/N) (∑nilgni) ]/[lgN- (1/N) (α (S-β) lgα+β (α+1) lg (α+1)) ]。式中, N为物种个体总数; ni为第i种的个体数; S为物种数; β为N被S整除以外的余数; α为(N-β) /S (彭少麟, 1996)。
3 结果与分析 3.1 群落组成变化茂兰喀斯特森林各种组主要树种如表 1。先锋种为阳性树种, 具光补偿点、饱和点高, 平均净光合速率高, 耐旱性强的特点; 次先锋种的光补偿点、饱和点较高, 平均净光合速率较高, 耐旱性较强; 过渡种的光补偿点较高, 平均净光合速率低, 具有一定的对弱光的利用能力, 耐旱性较弱; 次顶极种的光补偿点较低, 平均净光合速率较高, 耐荫性较强, 耐旱性较强; 顶极种为耐荫树种, 光补偿点低、饱和点高, 平均净光合速率中等, 耐旱性较强。
|
|

退化喀斯特群落自然恢复过程中, 各种组在群落中的优势地位发生更替(图 1), 在早期阶段, 先锋种占优势, 之后, 逐渐降低, 最终被淘汰, 而顶极种则刚好相反。次先锋种和过渡种变化相似, 在灌丛灌木阶段和灌乔过渡阶段占优势地位, 进入常绿落叶阔叶林阶段衰退, 而次先锋种衰退速度快于过渡种; 次顶极种在早期阶段变化较小, 从灌乔过渡阶段开始优势地位逐渐提高。各种组间的协变关系(Jhon, 1990)可反应一个种组变化对另一种组是抑制(负协变关系)或促进(正协变关系)作用。表 2表明:群落恢复过程中, 先锋种与其它种组呈负协变关系, 即相互抑制, 先锋种的衰退有利于其它种组地位提高, 特别是顶极种, 而顶极种重要性的提高有利于次顶极种发展却抑制了次先锋种、过渡种发展。因此, 种组变化是由先锋种, 经次先锋种、过渡种、次顶极种向顶极种逐渐替代。

|
图 1 群落组成、盖度、密度变化 Fig. 1 The change of community composition, coverage and density —◆—先锋种Pioneer species; —■—次先锋种Sub-pioneer species; —▲—过渡种Transition species; —■—次顶极种Sub-climax species; —●—顶极种Climax species. A—草本群落阶段Herbaceous stage; B—草灌群落阶段Herbaceous-scrub stage; C—灌丛灌木阶段Scrub-shrub stage; D—灌乔过渡阶段Shrub-tree stage; E—常绿落叶阔叶林阶段Evergreen and deciduous forest stage; F—顶极群落阶段Climax stage. |
|
|

随着退化群落恢复, 群落高度逐渐增加(表 3), 层次分化逐渐明显。草本群落阶段, 仅草本层一个层次, 树木个体与草本植物混生, 平均高0.6 m; 草灌过渡阶段开始出现灌木层, 但灌木高度低、个体小、盖度低; 灌丛灌木群落阶段, 形成典型的灌木层和草本层, 灌木层发育良好; 灌乔过渡阶段, 小乔木占据一定优势, 形成乔木层、灌木层和草本层, 但前两层分化不甚明显; 至常绿落叶阔叶林阶段后, 乔木层、灌木层、草本层分化明显, 在顶极群落中乔木层可达16.7 m, 多分化为两个亚层。
|
|
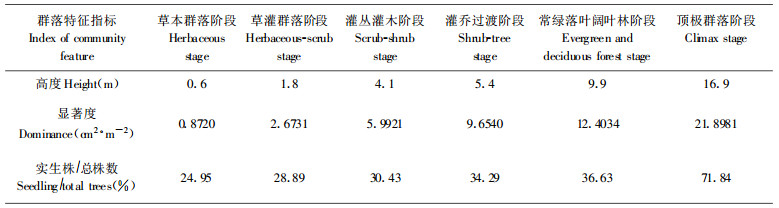
群落盖度随群落恢复早期阶段逐渐增大, 并在灌丛灌木阶段以后趋于稳定(图 1);显著度逐渐增大, 但在常绿落叶阔叶林阶段之前, 各阶段显著度增长速度相对较小, 之后增长迅速, 反应了在常绿落叶阔叶林阶段之前的群落恢复过程中以高生长为主, 之后为明显的粗生长。
群落主要层次密度变化体现了群落水平结构变化(图 1)。草本群落阶段, 林木个体幼小, 可容纳数量大, 密度迅速增加, 到草灌过渡阶段, 密度达到最大, 之后, 随着个体增大, 竞争分化剧烈, 密度迅速下降, 顶极群落阶段个体最大, 密度最小。
喀斯特森林退化后, 群落无性更新能力强, 草本群落阶段萌生株数可达总株数的75.05%, 随群落恢复, 萌生株数减少而实生株数增大, 顶极群落中实生株数可达71.84%。
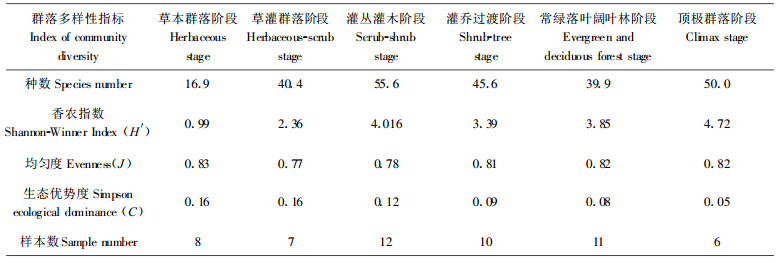
3.3 群落多样性变化退化群落自然恢复过程中, 其组织结构变化用多样性指数变化反应(表 4)。总体上看, 群落恢复趋势向组成物种增多, 多样性指数、均匀度指数上升而生态优势度降低的方向发展, 说明了退化群落的生境改善是向着顶极群落与喀斯特地貌双重作用形成的复杂多样的生境方向发展, 生境资源空间配置更趋复杂。早期阶段, 喀斯特生境干旱, 变化剧烈, 树种较少, 多样性指数低。群落恢复到灌丛灌木阶段, 早期严酷的生境状况得到改善, 大量物种侵入并生存, 物种骤增, 尽管个体数量分布不均匀, 均匀度不高, 但群落多样性指数迅速上升, 达到一个峰值, 之后至灌乔过渡阶段, 因种间竞争分化, 群落自然稀疏多样性下降, 之后又逐渐增大, 至顶极群落阶段为最高值。
|
|
随着退化群落自然恢复, 群落生物量逐渐积累增大, 至常绿落叶阔叶林阶段可恢复到顶极阶段的68.03% (表 5)。乔木层生物量在灌乔过渡阶段开始占优势并随之增加。灌木层生物量在草灌过渡阶段开始占优势, 至灌丛灌木阶段最高, 之后逐渐减小; 草本层生物量在草本群落阶段为优势, 随之逐渐减退。草本层的消退是群落走向成熟的标志之一。
|
|
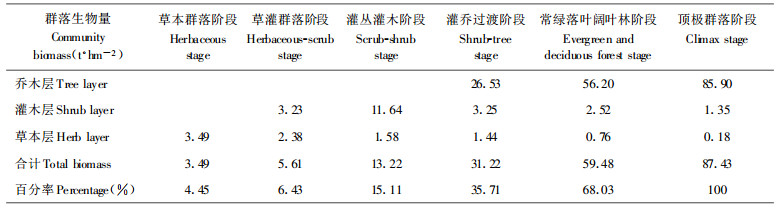
乔灌木生物量逐渐增大, 先锋种生物量最小而顶极种最大, 各种组生物量呈此消彼长的变化(表 6)。各阶段种组生物量的大小反应了其对群落恢复所作贡献大小。先锋种在早期阶段生物量占优势, 对早期群落结构建立, 功能的恢复起决定作用; 次先锋种、过渡种生物量在灌丛灌木阶段和灌乔过渡占优势, 对结构简单、功能低下的早期群落向结构功能复杂的顶极群落过渡起关键作用, 顶极种和次顶极种集中分布于常绿落叶阔叶林和顶极群落阶段, 对维护群落稳定和群落功能起重要作用。
|
|
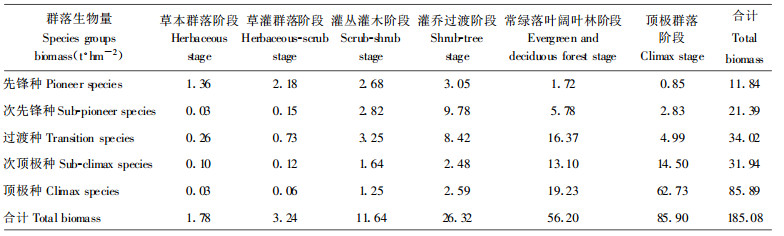
恢复过程中各种组生物量最大值出现的演替阶段不同(表 6), 先锋种、次先锋种、过渡种生物量最大值分别出现在草灌过渡阶段、灌乔过渡阶段和常绿落叶阔叶林阶段, 次顶极种与顶极种的最大值都在顶极群落阶段, 但顶极种明显占据优势, 这也反应了种组间的替代是从先锋种, 经次先锋种、过渡种至次顶极种和顶极种的过程。
4 小结退化喀斯特群落自然恢复过程中, 种组替代规律是先锋种经次先锋种、过渡种最终被次顶极种和顶极种所取代。退化喀斯特群落通过自然恢复总是向结构更复杂、更完善的方面发展, 体现于群落物种多样性、高度、盖度提高, 密度降低, 层次增多、分化明显, 显著度增大, 生物量积累增加。随群落演替萌生株数减少而实生株数增大, 群落更新对策由早期阶段无性更新向后期阶段有性更新发展。
杜道林, 刘玉成, 苏杰. 1996. 茂兰喀斯特山地广东松种群结构和动态初步研究. 植物生态学报, 20(2): 159-166. |
方任吉, 刘宗群.亚热带石灰岩山地存在常绿阔叶林———以南川金佛山为例.见钟章成编著, 常绿阔叶林生态学研究.重庆: 西南师范大学出版社, 1988, 537~549
|
郭魁士. 1940. 广西石灰岩区土壤之初步观察. 土壤, 1(4): 32-47. |
何道泉. 1993. 广东石灰岩地区的森林植物及其恢复问题. 热带地理, 13(3): 213-218. |
侯学煜. 1946. 贵州盘县之植物组合与土壤之初步观察. 土壤, 5(1): 53-61. |
侯学煜. 1952. 贵州省南部植物群落. 植物学报, 1(2): 65-106. |
胡舜士, 王献溥. 1982. 广西阳朔石灰岩同地乌冈栎的群落学特点及其在植被分类中的位置. 植物学报, 24(3): 264-272. |
梁士楚. 1992. 贵阳喀斯特山地云贵鹅耳枥种群动态研究. 生态学报, 12(1): 53-60. DOI:10.3321/j.issn:1000-0933.1992.01.013 |
彭少麟. 1996. 南亚热带森林群落动态学. 北京: 科学出版社, 84-99.
|
苏宗明. 1990. 广西石灰岩山地封山育林效果的分析. 广西植物, 10(4): 343-350. |
屠玉麟. 1989. 贵州喀斯特森林的初步研究. 中国岩溶, 8(4): 282-290. |
王献溥. 1981. 广西石灰岩地区常绿、落叶阔叶混交林的群落学特点. 东北林学院学报, (3): 30-47. |
吴春林. 1991. 广西热带石灰岩季节雨林分类与排序. 植物生态学与地植物学学报, 15(1): 17-25. |
肖育檀. 1988. 湖南石灰岩石山青冈栎落叶树混交林的群落分析. 中国林学院学报, 8(2): 146-154. |
徐樵利. 1993. 中国南方石灰岩荒山开发利用新探. 自然资源学报, 8(2): 115-121. DOI:10.3321/j.issn:1000-3037.1993.02.003 |
杨继镐, 汪炳根, 唐俊. 1990. 广西大青山石灰岩山地土壤理化性质的演替及其造林绿化. 林业科学, 26(5): 402-409. |
杨汉奎, 程任泽. 1991. 贵州茂兰喀斯物森林群落生物量研究. 生态学报, 11(4): 307-312. DOI:10.3321/j.issn:1000-0933.1991.04.010 |
喻理飞, 朱守谦, 魏鲁明. 1998a. 贵州喀斯特台原亮叶水青冈种多度结构研究. 山地农业生物学报, 17(1): 9-15. |
喻理飞, 朱守谦, 叶镜中. 1998b. 退化喀斯特群落自然恢复过程研究———自然恢复演替系列. 山地农业生物学报, 17(2): 71-77. |
喻理飞, 朱守谦, 叶镜中. 2000. 退化喀斯特森林自然恢复评价研究. 林业科学, 36(6): 12-19. DOI:10.3321/j.issn:1001-7488.2000.06.002 |
张祝平, 何道泉, 敖惠修. 1993. 粤北石灰岩山地主要造林树种的生理生态学特性. 植物生态学与地植物学学报, 17(2): 133-142. |
周政贤主编.茂兰喀斯特森林科学考察集.贵阳: 贵州科技出版社, 1987: 1~23
|
朱守谦主编.喀斯特森林生态研究(Ⅰ).贵阳: 贵州科技出版社, 1993: 1~73
|
朱守谦主编.喀斯特森林生态研究(Ⅱ).贵阳: 贵州人民出版社, 1997: 9~47, 55~107, 128~159
|
朱守谦, 魏鲁明, 陈正仁. 1995. 茂兰喀斯特森林生物量构成初步研究. 植物生态学报, (4): 354-367. |
D. Muller-Dombois H. Ellenberg著. (鲍显诚等译).植被生态学的目的和方法, 北京: 科学出版社, 1986: 247~252
|
Jhon A.拉德维格, James F.蓝诺兹著. (李育中等译).统计生态学———方法和计算入门.呼和浩特: 内蒙古大学出版社, 1990, 94~100
|