 2001, Vol. 37
2001, Vol. 37文章信息
- 何东进, 洪伟, 吴承祯, 蓝斌.
- He Dongjin, Hong Wei, Wu Chengzhen, Lan Bin.
- 毛竹林生态系统能流经济阈值模拟研究
- A STUDY ON THE SIMULATION OF ECONOMICAL THRESHOLD OF ENERGY FLOW IN PHYLLOSTACHYS PUBESCENS ECOSYSTEM
- 林业科学, 2001, 37(专刊1): 194-199.
- Scientia Silvae Sinicae, 2001, 37(专刊1): 194-199.
-
文章历史
- 收稿日期:2001-01-08
-
作者相关文章


2. 鹭江大学 厦门 361000
2. Lujiang University Xiamen 361000
经济阈值(Economical threshold, 简称ET)首先由Stern提出(Stern, 1959), 它是现代害虫防治的基本决策依据。经济阈值的概念与方法目前在许多领域得到应用与发展(Headley, 1972;盛承发, 1985;1989;苏祥瑶等, 1987;夏乃斌等, 1992;1993), 其发展趋势之一是研究多种害虫的复合经济阈值及其替代指标(盛承发, 1989)。生态系统的能流分析是系统功能研究的重大课题, 如何将能流分析与复合经济阈值联系起来, 是生态系统研究中的重要内容(竺锡武等, 1994)。
毛竹(Phyllostachys pubescens)是最重要的一种竹类资源, 具有生长快、产量高、用途广和可持续更新等特点, 毛竹林的经营开发已成为山区农民脱贫致富的重要途径之一。国内有不少学者致力于毛竹丰产栽培、生物量、能量的分配及不同类型生态系统之间的能流比较等方面的研究(周芳纯, 1986;洪伟等, 1988, 1998a, 1998b;蓝斌等, 2000;何东进等, 1999, 2000a, 2000b)。但是, 毛竹的病虫害常使其产量遭受巨大的损失, 目前在毛竹林经营地区所制定的“防治指标”多数是“经验指标”或“人为指标”, 存在很大的局限性与任意性, 因此, 如何从能量生态学角度出发, 运用经济阈值手段, 将能流分析与经济阈值结合起来, 科学制定毛竹林的优化管理政策是山地毛竹综合开发中亟待解决的问题, 而前人在这方面未曾有过报道。有鉴于此, 笔者在毛竹标准地调查材料的基础上, 试图对毛竹林生态系统能流经济阈值进行研究, 为毛竹林经营的丰产、稳产与优化管理提供理论依据。
1 研究区概况与材料收集 1.1 研究区概况研究区设在福建省建瓯市.建瓯市位于福建省北部, 地理坐标26°14′~ 28°20′N, 117° 02′~ 119°07′E之间, 海拔在68 ~ 1822.2 m, 年平均温度在17.5 ~ 19.3 ℃之间, 极端最高气温41.4 ℃, 最低气温-9.5 ℃, ≥10 ℃的年活动积温在5000 ℃以上, 年平均降水量在1604 ~ 2399 mm之间, 年平均相对湿度为80 %。土壤类型以红壤为主, 其次是黄壤, 土层厚度一般为80 ~ 150 cm, 土壤质地结构较好, 多为轻壤、中壤, 少数为砂壤和重壤, pH值4.9 ~ 5.4。
调查样地设在建瓯市房道镇房道村的毛竹示范林中, 海拔在300 m左右, 坡度在20°左右, 一般为中、下坡, 群落总盖度50 %~ 70 %, 立木层为单一的毛竹, 层盖度为60 %~ 85 %, 林下以草本植物为主, 平均高度为5 ~ 15 cm, 主要有凌霄(Campsis grandiflora)、悬钩子(Rubus palmatus)、尖叶菝葜(Smilax arisanensis)、芒萁(Dicroanopteris dichotoma)、五节芒(Miscanthus floridulus)、黑莎草(Gahnia tristis)、地稔(Melastoma dodecandrum)、沿海紫荆牛(Ardisia punctata)、江南卷柏(Selaginella moellendorffii)等, 林木、蕨类(Pteridophyta sp.)植物较少。
1.2 样地调查方法用样方法在丰产毛竹林中对毛竹进行本底调查, 调查面积为10个10 m ×10 m的样方, 记录样方内每株立竹的高度、枝下高、胸径和年龄。灌木层、草本层则在样方内随机选取5 m ×5 m的小样方2个, 记录每种灌木、草本和藤本植物的高度、盖度、株数, 同时记录竹林的季相及土壤情况。地下部分在样方内以选定的标准竹为中心, 选取1 m ×1 m的小样方2个, 记录竹鞭、竹蔸、竹根、其它根及土壤剖面情况。
1.3 样品采取、生物量与热值测定在样方内以平均胸径为基准, 参考平均年龄选伐标准竹9株, 以根茎交界处为分界线, 其上为地上部分, 其下为地下部分。地上部分以1 m为区分段, 测定竹秆、竹枝、竹叶的鲜重, 并分别取样带回, 在105 ℃下烘干至恒重, 然后换算成标准地生物量; 地下部分以10 cm为区分段分层分竹鞭、竹根、竹蔸和其它植物根, 用水冲洗去泥后, 阴干称其鲜重, 并取样烘干称重, 磨碎过筛待测。林下植物以样竹为中心, 作1 m ×1 m的样方, 割取地上部分称其鲜重, 作同上处理。
用GR-3500型氧弹式量热计(Hughes, 1971), 在室温20 ℃对待测样品进行热值测定, 每个样品重复2 ~ 3次。
2 毛竹林生态系统能流经济阈值的概念与研究方法 2.1 毛竹林生态系统能流经济阈值的概念竺锡武等在研究棉田生态系统能流经济阈值时, 将系统能流经济阈值定义为:作物被害虫摄食的某一能量, 达此值时应立即采取控制措施, 否则将引起等于这一措施期望代价的期望损失(竺锡武等, 1994)。系统能流经济阈值(Energy economical threshold, 简记EET)是指研究系统可以向害虫亚系统输出能量的最大允许临界值。因此, 笔者参照竺锡武等所给出的棉田生态系统能流经济阈值定义, 将毛竹林生态系统能流经济阈值定义为:“毛竹林被害虫摄食的某一能量, 达到此值时应立即采取防治措施, 否则将引起等于这一措施期望代价的期望损失。”
2.2 研究方法根据“挽回损失=措施代价”的基本原则(盛承发, 1985), 研究毛竹林生态系统能流经济阈值。
(1) 建立毛竹林生态系统能流动态模型 分室模型(Compartment Model, Smith, 1970)是目前生态学中用得较多的一种模型, 在生态系统能量研究中广泛应用的能流分析方法(陆国泉等, 1990;黄大明等, 1992;何东进等, 2000c, 2000d)。本文运用分室理论建立毛竹林生态系统能流动态模型。
(2) 确定措施代价 假设措施代价是毛竹林未受害时经济产量的某一数值。这里所言的毛竹林经济产量(或毛竹林产量)系指毛竹林的能量。本文以人工模拟去叶、去枝作为病虫害对毛竹林造成的危害来研究毛竹林生态系统能流经济阈值。因此, 措施代价为某一设定值, 不失一般性, 本文假设措施代价为2%。
(3) 建立能流经济阈值模型 利用计算机对系统能流动态模型中参数进行模拟, 求得能流经济阈值。对系统模型中的害虫能量参数a32(表示去叶)和a12(表示去枝)分别进行模拟, 找出当毛竹林产量(能量)值等于a32或a12时毛竹经济产量的98%(原设定防治代价为2%)时的阈值参数a32和a12, 就可以得到毛竹林生态系统能量经济阈值(EET), 即经济阈值模型。
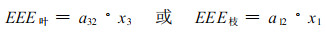
|
(1) |
式中, EEE叶、EET枝分别表示以去叶或去枝模拟害虫为害的能流经济阈值, x3、x1分别表示系统中叶、枝的能量现存量。
(4) 毛竹林补偿与超补偿效应通过建立毛竹林生物量与去叶率、去枝率的关系模型, 分析其反应曲线, 确定毛竹林的补偿点与超补偿点。
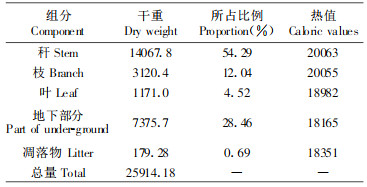
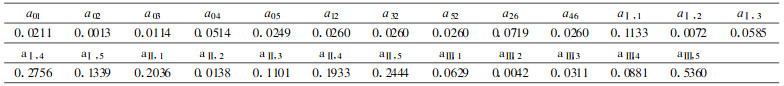
3 结果与分析 3.1 毛竹各组分样品干重、热值及生物量毛竹各组分样品干重、热值的测定是计算毛竹林生物量与群落能量动态变化的基础。现将各组分样品干重、热值的测定结果列于表 1。
|
|
在毛竹林生态系统中, 毛竹各组分间的能量随时间的动态变化而变化。本文以1995年在建瓯市房道镇毛竹丰产林的调查及测定分析资料为依据, 将系统划分为竹秆、竹枝、竹叶、地下部分及凋落物5个子系统, 并结合时间的动态变化建立图 1所示的能流动态模型, 其中系统外能量变化仅考虑新竹和土壤。

|
图 1 毛竹林生态系统能流动态模型 Fig. 1 The dynamic model of energy flow of Phyllostachys pubescens ecosystem ①新竹New bamboo; 竹枝Branch; 竹秆Stem; 竹叶Leaf; 凋落物Litter; 地下部分Part of under-ground; 土壤Soil; Ⅰ度竹1-year-old Bamboo; Ⅱ度竹2~ 3-year-old Bamboo; Ⅲ度竹4~ 5-year-old Bamboo; Ⅳ度竹6~ 7-year-old Bamboo; Ⅴ度竹8~ 9-year-old Bamboo. |
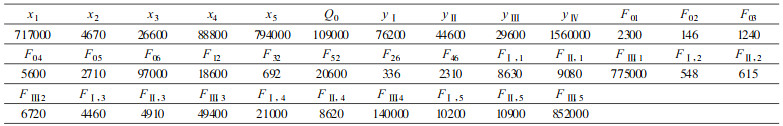
图 1是毛竹林生态系统能量流动态分室模型。其中能量源和能量汇用旗表示, 暂时的能量积累分室用长方形表示, 时间动态分室用圆圈表示, 直接的能源用带箭头的实线表示。x表示状态变量(kJ· m-2), y表示时间分室状态变量(kJ·m-2), 积累在不同分室的数量表示为:(1)竹枝x1;(2)凋落物x2; (3)竹叶x3;(4)地下部分x4;(5)竹秆x5;(6)yi(i=Ⅰ ~ Ⅳ)表示第i度竹。各分室能量以Ⅱ、Ⅲ、Ⅳ度竹能量之和作为动态考虑。
|
|
根据该分室模型, 就可以得到相应的毛竹林各分室能量流动一阶微分方程组模型:
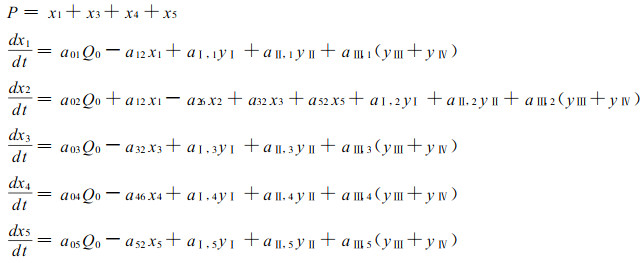
|
(2) |
式中, P为系统的最终生物量或能量(kJ·m-2); aij(=Fij/xi)为从i分室流向j分室的能通率参数; Fij为毛竹能量从i分室流入j分室的流通量; xi为各组分的状态变量(kJ·m-2), xi(i =1 ~ 5)分别表示毛竹的枝、凋落物、叶、地下部分及秆; yj为第j度竹(j =Ⅰ ~ Ⅳ); Q0为新竹状态变量(kJ·m-2)。各参数值见表 3。
|
|
由于调查林分属毛竹丰产林, 极少受到病虫害的危害, 将其视为未受害毛竹林系统, 这时a32=0.0260, a12=0.0260, 从而求得毛竹林的最终产量(以能量表示)为:
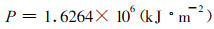
|
(3) |
当毛竹受病虫害危害(这里采用人工模拟去叶、去枝的方法)时, 根据假设其措施代价为2%, 则可求出毛竹的期望最终产出为:

|
(4) |
现以P′为准, 分别对a32(去叶)和a12(去枝)进行参数模拟, 所得的模拟参数阈值分别为:
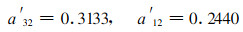
|
(5) |
于是得到毛竹林生态系统能流经济阈值为:

|
(6) |
式中, x3、x1分别为竹叶、竹枝的现存量。
3.4 毛竹补偿与超补偿效应毛竹受害后的损失, 不仅与危害毛竹林的害虫种群密度, 而且也与毛竹林对虫害所做出的反应有关。国内、外研究表明, 补偿与超补偿效应广泛存在于自然界的许多植物中(Bank, 1976;盛承发, 1989;夏乃斌等, 1993), 为进一步探讨毛竹的补偿与超补偿效应, 分别建立毛竹经济产量损失率(y)与去叶率(x枝)、去枝率(x叶)的关系模型, 并绘制其关系曲线图(图 2):

|
(7) |
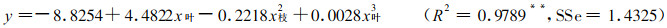
|
(8) |

|
图 2 毛竹林生态系统能量损失率与去叶率(去枝率)的关系 Fig. 2 Relationship curves between the loss of system energy and defoliation(branch-cut)rate ---◆---去枝率Branch-cut rate; —■—去叶率Defoliation rate. |
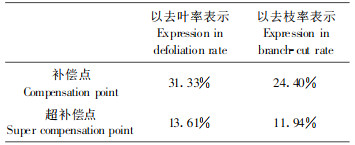
由图 2可以看出, 毛竹在受到病虫害危害时, 产量并没有立即减少, 只有危害超过一定程度时, 其产量才会下降, 可见毛竹存在着补偿与超补偿效应。毛竹的补偿点约为去叶率31%或去枝率24%左右, 其超补偿点约为补偿点的一半, 分别为去叶率14 %或去枝率12%左右。为了精确计算出毛竹的补偿点与超补偿点, 令式(7)、(8)中y=0及一阶导数y′=0, 便可求出补偿点与超补偿点, 结果列于表 4。
|
|
害虫的摄食能量损耗随着害虫的种类、种群密度、为害时间的不同而不同, 用能量指标代替种群密度或多种群复合指标, 不仅可以将不同害虫的取食为害程度用能量统一起来, 而且可以将害虫为害的瞬时影响和对作物经济产量的最终影响联系起来, 因此具有综合的意义。
本文以去叶(去枝)模拟病虫害对毛竹林产生的危害, 将能流分析与经济阈值相结合, 对毛竹林生态系统能流经济阈值进行研究, 结果表明:所求的阈值参数分别为a′32=0.3133和a′12=0.2440, 说明当毛竹林受病虫害危害, 在去叶率(或去枝率)不超过31.13%(或24.40%)时, 毛竹林的经济产量并未受到损失, 不必进行防治, 当超过该临界值时必须进行防治, 否则将引起毛竹产量的损失, 从而为实际指导毛竹林经营提供依据; 从能流阈值模型可以看出, 当a′32和a′12保持不变时, 毛竹能流经济阈值(EET)随着竹叶、竹枝现存量的增大而增大, 说明毛竹的耐害能力随着毛竹的生长变化而变化, 这与前人在其它领域研究有着相类似的结果(竺锡武等, 1994);毛竹与农作物(棉花、蕃茄、豆类、果树和粮食植物等)及森林植物(油松)等一样存在着补偿与超补偿效应, 其补偿点分别为去叶率31.33%或去枝率24.40%, 超补偿点约为补偿点的一半, 分别为去叶率13.61%或去枝率11.94%。
EET值的精确与否, 很大程度上依赖于系统能流模型的模拟精度, 本文立足于毛竹结构能量动态特征, 结合时间动态, 利用试验分析方法建立系统能流动态模型, 考虑的因素尚未全面, 尤其未考虑林下植被的能流问题, 因此, 所建的模型不够完善。植物的补偿作用实质是能量分配的一种优化表现, 与植物内部复杂的机理因素及外部的营林措施密切相关, 本文给出的毛竹的补偿点与超补偿点仅是从模型出发确定的, 因此, 比较粗糙而不够理想, 这些有待于今后进一步探讨改进。
洪伟, 杨居暖. 1988. 毛竹产量新模型研究. 竹子研究汇刊, 7(1): 1-12. |
洪伟, 郑郁善, 邱尔发. 1998a. 毛竹丰产林密度效应研究. 林业科学, 34(专): 1-5. |
洪伟, 兰斌, 吴承祯, 等. 1998b. 毛竹林能量分配的研究. 林业科学, 34(专): 78-81. |
黄大明, 赵松岭. 1992. 矮嵩草草甸能量动态的分室模型研究. 生态学报, 12(2): 119-124. DOI:10.3321/j.issn:1000-0933.1992.02.007 |
何东进, 洪伟, 吴承祯, 等. 1999. 毛竹种群光能利用率的研究. 福建林学院学报, 19(4): 324-326. DOI:10.3969/j.issn.1001-389X.1999.04.011 |
何东进, 洪伟, 吴承祯, 等. 2000a. 毛竹林林分平均胸径模型预测模型的研究. 林业科学, 36(专刊1): 148-153. |
何东进, 洪伟, 吴承祯. 2000b. 毛竹林各组分能量估算模型的研究. 应用与环境生物学报, 6(5): 412-418. |
何东进, 洪伟, 吴承祯, 等. 2000c. 毛竹林生态系统能量动态规律的研究. 竹子研究汇刊, 19(1): 63-67. |
何东进, 洪伟, 吴承祯, 等. 2000d. 毛竹林生态系统能量动态分室新模型. 植物资源与环境学报, 9(4): 9-13. |
蓝斌, 何东进, 洪伟, 等. 2000. 闽北毛竹林生态系统能量分配规律的研究. 应用生态学报, 11(2): 193-195. DOI:10.3321/j.issn:1001-9332.2000.02.009 |
陆国泉, 张树源, 杨福囤, 等. 1990. 矮嵩草草甸生长季节生产者亚系统分室模型. 植物生态与地植物学学报, 14(2): 97-102. |
盛承发. 1985. 华北棉区第二代棉铃虫的经济阈值. 昆虫学报, 28(4): 382-389. |
盛承发. 1989. 害虫经济阈值的研究进展. 昆虫学报, 32(4): 492-499. DOI:10.3321/j.issn:0454-6296.1989.04.001 |
苏祥瑶, 林昌善. 1987. 粘虫危害小麦的动态经济阈值的研究. 生态学报, 7(4): 322-329. |
夏乃斌, 屠泉洪, 宋长义, 等. 1992. 油松毛虫危害油松的动态经济阈值的研究. 北京林业大学学报, 14(4): 9-19. |
夏乃斌, 屠泉洪, 宋长义, 等. 1993. 油松林对油松毛虫危害的补偿与超补偿效应的研究. 生态学报, 13(2): 121-129. DOI:10.3321/j.issn:1000-0933.1993.02.014 |
竺锡武, 汪世泽. 1994. 棉田生态系统能流经济阈值的初步研究. 生态学报, 14(1): 63-67. |
周芳纯. 1986. 毛竹林丰产技术研究总结报告. 竹类研究, 5(3): 1-39. |
Bank C J. 1976. Effect of Aphis fabae Scop.and of its attendant ants insect predators on yields of field beans(Victa faba). Ann.Appl.Biol, 60: 445-453. |
Headley J C.Defining the economic threshold in Pest Control Strategies for the Future.Nas, Washington D.C., 1972, 100~108 http://cn.bing.com/academic/profile?id=fb5a52bbbafb449973dad2215d82c146&encoded=0&v=paper_preview&mkt=zh-cn
|
Hughes M K. 1971. Seasonal calorific values from a deciduous woodland in England. Ecol, 52: 923-926. DOI:10.2307/1936045 |
Stern V M. 1959. The integration of chemical and biological control of the spotted aphid. Hilgarolia, 29(2): 81-101. DOI:10.3733/hilg.v29n02p081 |