 2001, Vol. 37
2001, Vol. 37文章信息
- 刘健, 陈平留.
- Liu Jian, Chen Pingliu.
- 建阳武夷山区米槠群落特征的初步研究
- STUDY ON THE COMMUNITY CHARACTERISTIC OF CASTANOPSIS CARLESII FOREST IN WUYI MOUNTAINOUS REGION OF JIANYAN, FUJIAN
- 林业科学, 2001, 37(专刊1): 173-176.
- Scientia Silvae Sinicae, 2001, 37(专刊1): 173-176.
-
文章历史
- 收稿日期:2001-01-08
-
作者相关文章


米槠(Castanopsis carlesii)林是我国东部湿润亚热带地区常绿阔叶林中重要群系之一(章浩白, 1993)。对其群落特征、种群间的相互关系以及演替趋势系统深入的研究, 为实施天然林保护工程, 提高森林经营水平, 模拟自然群落类型, 改善景观的异质性格局, 评价森林群落或生态系统的状态, 具有重要的理论和现实意义。
1 数据的收集资料收集于建阳市焦溪林业采育场。在天然米槠林群落内典型选样设置8个面积为20 m×20 m的标准地, 每块标准地分成16个5 m×5 m样方。在各样方内, 对胸径(DBH) ≥5.0 cm以上的所有树木进行每木检尺, 测定其树种、胸径、树高、枝下高和冠幅, 实地绘制树冠投影图, 并记载土壤类型、海拔、坡向、坡位、坡度、土层厚度等。同时, 在每一标准地的4个角和中央设置2 m×2 m样方, 调查灌木、幼树、幼苗的种类、株数和苗(树)高。把胸径(DBH) ≥5.0 cm, 高度(H) ≥5.0 m的林木作为乔木层个体, 2 m≤H < 5.0 m的林木为灌木层个体, H < 2.0 m为下木层, 下木层包括灌木层和草本层。
2 数据处理群落物种多样性采用Simpson指数(D), Shannon-Wiener指数(H)和Shannon-Wiener均匀度(JH)测定(马克平等, 1994; 刘健, 1996)。
种群空间格局采用聚块性指数(m*/m) (Pielou, 1988)。
群落种群间的总体联结关系检验采用方差比率(RV)法(Schluter, 1984; 刘健等, 1999)。
种群生态位采用Shannon-Wiener指数(Colwell, 1971)。
3 结果 3.1 群落结构根据调查结果, 标准地内共有维管植物65种, 隶属于30个科, 45个属。其中裸子植物2种, 双子叶植物58种, 单子叶植物2种, 蕨类3种。群落外貌呈深绿, 郁闭度达0.85。从表 1看出, 乔木层中米槠的优势最为突出, 其重要值百分率为32.77%, 是该群落的第1优势种和建群种。南岭栲(Castanopsis fordiii)的重要值百分率为10.76%, 木荷(Schima superba)重要值百分率为9.72%, 栲树(Castanopsis fargesii)重要值百分率为8.99%, 它们均为第2优势种和共建种。其它种类的重要值百分率均小于5%。应当看到, 一些灌木生长型种类的高大个体进入乔木层, 不仅丰富了该层成份, 也反映出这些种类对特殊生境条件的生态适应性, 从而增加了乔木层结构的复杂性, 这是该群落结构上的一大特点。群落内部结构分化明显, 可划出5个垂直结构层次(米槠群落垂直结构剖面图略)。
|
|
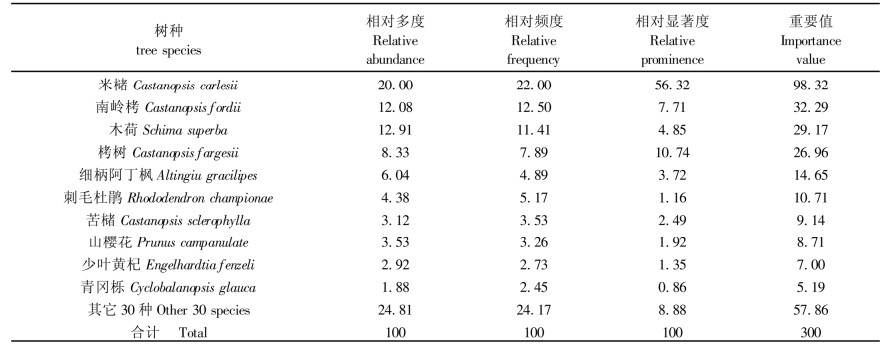
乔木层可分为3亚层, 第1亚乔木层高16~25 m, 平均高为19.1 m, 主要由米槠、南岭栲和苦槠(Castanopsis sclerophylla)等树木构成, 郁闭度在0.4左右; 第2亚乔木层高11~15 m, 平均高12.8 m, 南岭栲、木荷、栲树、米槠等繁殖层的多数个体分布其中, 常见种类还有细柄阿丁枫(Altingiu gracilipes)、苦槠、山樱花(Prunus campanulate)、猴欢喜(Sloanea sinensis)、青冈栎(Cyclobalanopsis glauca)等, 该亚层密度较大, 种类较多, 郁闭度近0.6;第3亚乔木层高度在5~10 m, 由于一些乔木幼树和大灌木进入, 成分较复杂, 主要有刺毛杜鹃(Rhododendron championae)、密花山矾(Symplocos congesta)、树参(Dendropanax dentiger)、厚壳桂(Cryptocarya chinensis)、华南桂(Cinnamomum austrosinensis)、刨花楠(Machilus pauhoi)、山杜英(Elaeocarpus sylvestris)、木(Loropetalum chinense)、木荚红豆(Ormosia xylocarpa)等以及上层乔木树种。因受上层树冠遮蔽, 分布较零散, 郁闭度仅约0.2。
灌木层植物种类以壳斗科、茶科和山矾科为主, 更新苗以南岭栲、木荷、米槠为主, 灌木植物有老鼠刺(Itea chiensis)、狗骨柴(Tricalysia dubia)等。
草本层通常分布不连续, 多集中于林缘和林窗下, 种类较丰富, 达53种, 但在不同层片和小群落环境中优势种各异。
3.2 群落物种多样性分析在森林群落自然演替过程中, 逐步趋向相对稳定的气候顶极群落, 其各层次的多样性指数值趋向一致(彭少麟, 1996)。从表 2中可以看到Shannon-Wiener指数为2.5~5.0, Shannon-Wiener均匀度指标为0.60~0.87左右, 且各层次的物种多样性指数表现出相对一致, 表明该群落是相对稳定的。特别值得注意的是Shannon-Wiener指数和Shannon-Wiener均匀度指标明显高于中亚热带常绿阔叶林(刘健, 1996), 而与一些南亚热带常绿阔叶林接近(彭少麟, 1996)。
|
|

采用方差比率(RV)法计算得RV=1.0381>1, W=N·RV=128×1.0381=132.8768, 由于χ20.95, 128=102.87 < W < χ20.05, 128=155.40, 故群落乔木层40个种群间的整体联结关系表现为不显著的正联结。结果表明米槠树种生长茂盛, 其它阔叶树种发育已较滞止。究其原因, 所调查的林分, 以米槠为优势树种, 该树种具有耐庇荫, 喜温暖湿润的特点, 枯枝落叶在林地上分解快, 形成的腐殖质层厚, 是改变土壤结构和提高土壤肥力的树种。且在荫蔽环境中, 种子能正常发芽, 生长很快, 从而在与其它树种的竞争中, 常处于优势地位, 这在文献(章浩白, 1993)得以证实。另外林层结构上, 米槠占据了第1亚乔木层总株数的54%, 南岭栲和栲树仅分别占该层总株数的8.6%和11.4%, 其它各层中建群种和共建种具有较均匀的分布, 各种群之间能占据的空间生态位不同, 彼此之间能协调地发展。此外, 从群落物种多样性分析已证实, 该群落是相对稳定的, 因而反映在多物种的种间联结上是不显著的正联结。
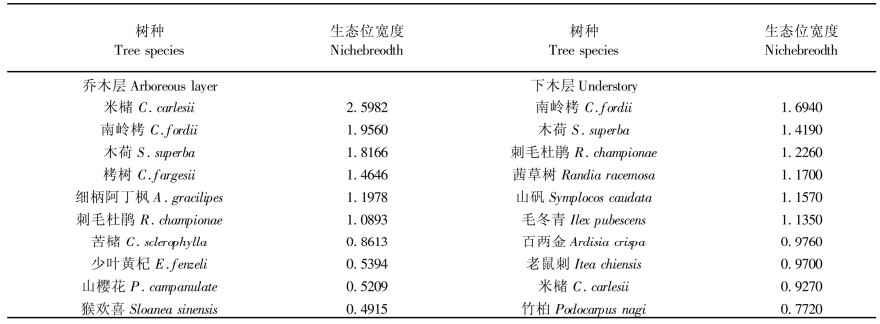
3.4 种群生态位种群的空间生态位是种群对生境因素的适应以及利用资源多少的结果。同一时间里观察不同种群在同域里的生态位变化, 应该反映出生境对种群的影响和种间竞争特性。从表 3看出, 乔木层米槠的生态位宽度值最大, 南岭栲和木荷生态位宽度在1.8~2.0之间十分相近; 在下木层中南岭栲和木荷的生态位宽度位居第1、2位, 米槠的生态位宽度落居第9位, 这表明随着第1亚乔木层中米槠种群个体自然成熟死亡, 在种群对光资源的利用性竞争中, 阳性阔叶种群的南岭栲和木荷占据优势, 从下木层至乔木层生态位宽度逐渐增大, 可预示米槠群落的演替进程有被日益增大的环境资源利用范围的南岭栲和木荷种群所取代的趋势。因而推论该群落目前的变化过程属进展演替类型是成立的。
|
|
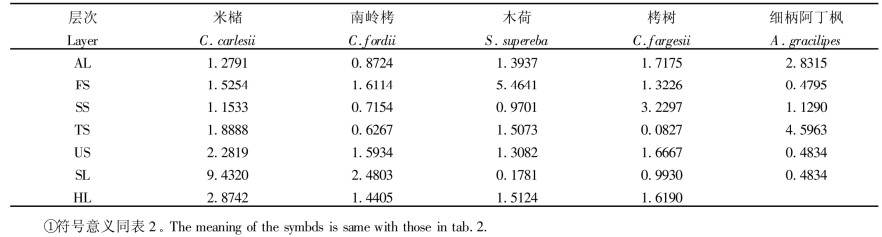
聚块性指数能表征种群中个体的聚集和扩散趋势, 指出种群个体空间变化的方向, 即若聚块性指数由大变小, 表明种群从这一阶段向下一阶段呈扩散的趋势; 若聚块性指数由小变大, 则表示种群呈聚集的趋势。从表 4可以看出米槠群落主要树种不同发育阶段呈现的空间格局变化趋势:米槠呈聚集分布, 表现为一种轻微的扩散趋势; 南岭栲的下木层呈聚集分布, 乔木层呈均匀分布, 表现出强烈的扩散趋势; 木荷和栲树呈聚集分布, 表现微弱的种群聚集趋势。而细柄阿丁枫的下木层呈均匀分布, 乔木层呈较强的聚集分布, 表现强烈的聚集趋势。以上这些种群空间格局变化趋势, 究其原因可能与树种喜光和萌芽能力大小有关。此外, 在第1亚乔木层中除细柄阿丁枫外均呈聚集分布。草本层中, 建群种、优势种和共建种的幼树、幼苗多, 从母树周围长出, 故具较强的集群分布特性。综上结果符合亚热带常绿阔叶林群落种群格局的典型特征(章浩白, 1993; 彭少麟, 1996)。
|
|
根据该群落的结构, 乔木层优势种的组合特征和种群结构特点, 把该群落命名为“米槠群落”是恰当的。
该群落的物种多样性指数明显高于中亚热带常绿阔叶林, 而与一些南亚热带绿阔叶林接近。表明它是中亚热带常绿阔叶林向南亚热带阔叶林过渡的水平地带性的类群之一。
群落各林层物种多样性指数趋向一致, 说明该群落是相对稳定的。
群落分林层的种群生态位指标的分析表明:可能经过相当时期后, 由于优势种群间的竞争, 在一定程度上影响群落的稳定性, 使目前处于次优势种的南岭栲和木荷等种群, 以其丰富的幼苗(树)贮备, 合理的年龄结构分布, 取代米槠的地位而成为第1优势种群。同时推论, 该群落目前的变化过程属进展演替类型。
在本群落中, 米槠处于旺盛生长时期, 且林下有一定的幼苗、幼树, 具有较强的天然更新能力。同时, 南岭栲和木荷也有一定的潜力, 因此, 有必要对林分进行抚育, 保护米槠幼苗、幼树, 改善其生长环境, 从而有利于米槠群落的发展。
刘健, 等. 1999. 天然针阔混交林优势种群间联结关系. 应用与环境生物学报, 5(5): 468-472. DOI:10.3321/j.issn:1006-687X.1999.05.007 |
刘健. 1996. 南平部分森林群落物种多样性分析. 福建林学院学报, 16(4): 324-328. |
马克平, 刘玉明. 1994. 生物群落多样性的测定方法Ⅰ, α多样性的测度方法(下). 生物多样性, 2(4): 231-239. DOI:10.3321/j.issn:1005-0094.1994.04.009 |
彭少麟. 1996. 南亚热带森林群落动态学. 北京: 科学出版社, 83-197.
|
章浩白. 1993. 福建森林. 北京: 中国林业出版社, 83-170.
|
Pielou E C(卢泽愚译).数学生态学(第二版).北京: 科学出版社, 1988, 308~331
|
Colwell R K. 1971. On the measurement of niche breadth and overlap. Ecology, 52: 567-576. DOI:10.2307/1934144 |
Schluter D. 1984. Avariance test for detecting species association, with some example applications, with some example applications. Echology, 65: 998-1005. |