 2001, Vol. 37
2001, Vol. 37文章信息
- 邱尔发, 洪伟, 郑郁善, 陈卓梅, 陈礼光.
- Qiu Erfa, Hong Wei, Zhen Yushang, Cheng Zhuomei, Cheng Liguang.
- 麻竹山地笋用林笋期叶片光合及呼吸性状研究
- STUDY ON THE PHOTOSYNTHETIC AND RESPIRATORY CHARACTER OF LEAVES OF FOREST PLANTED ON MOUNTAIN AND USED FOR SHOOT OF DENDROCALAMUS LATIFLORUS DURING GROWING SHOOTS
- 林业科学, 2001, 37(专刊1): 148-153.
- Scientia Silvae Sinicae, 2001, 37(专刊1): 148-153.
-
文章历史
- 收稿日期:2001-01-08
-
作者相关文章


麻竹(Dendrocalamus latiflorus)经济价值高、用途广泛、易栽培、周期短、且适应范围广, 是南亚热带、热带及部分中亚热带的重要经济竹种。近几年, 随着人们生活水平的提高, 健康意识的增强, 绿色食品脱颖而出。而麻竹笋作为天然的绿色保健食品, 味道鲜美, 营养丰富, 含有20种氨基酸和多种人体必需的微量元素, 且有清凉解暑的功效, 有素食第一之美称, 受到人们广泛的青睐(黄克福, 1992); 其各种笋罐头及笋干等系列产品, 畅销国内外市场; 除竹笋外, 其竹材及枝叶除传统的用途; 还可加工成竹工艺品、造纸等, 竹叶出口东南亚等国作为绿色食品的包装品; 另外, 麻竹根系发达, 具有较强的涵养水源, 防风保土的作用。过去, 麻竹只有在河边和房前屋后零散种植, 没有得到重视, 没有集约经营, 但随着人们对麻竹产品需求的不断增加, 过去的种植面积已远远不能满足人们的需求, 人们开始寻求更广阔的栽培空间; 另一方面, 由于近几年木材市场疲软, 林业不景气, 更换造林树种, 发展高效林业已成为走出困境的必然。麻竹笋期长, 产量高, 市场前景好, 经济价值高, 理所当然成为山地种植的探索竹种。经过几年的摸索, 山地栽培麻竹丰产技术正不断的改进, 竹农栽种麻竹的热情很高, 竹林面积正以迅猛的速度发展。
但是, 由于麻竹的经济价值近几年才开始被逐渐认识, 人们对山地麻竹栽培的认识主要还停留在感性认识上, 其生物及生态学特性也尚未被充分认识, 有关麻竹山地栽培特性研究文献颇少(周本智等, 1999; 肖贤坦等, 1996; 梁鸿木等, 1998)而有关麻竹光合及呼吸特性的研究目前尚未见报导, 制约山地麻竹栽培进一步向高效、丰产、优质、稳定、持续的方向发展, 有鉴于此, 研究山地麻竹光合及呼吸特性, 揭示麻竹生理基础, 这无疑对山地麻竹丰产经营实践和理论研究都有现实意义。
1 试验地自然概况及材料试验地设在国有南靖林场的炭坑工区, 位于东经117°21' ,北纬24°31' ,海拔280 m, 属闽南低山丘陵地带。年平均温度为21℃, 7月平均气温28℃, 10℃以上的活动积温7476℃, 终年无雪, 霜期短; 该地区气候温暖多雨, 年降雨量1720 mm, 雨量分布不均, 较集中于春夏两季; 年日照时数2052 h, 平均年无霜期320 d, 植被以沙岩土质发育的山地红壤为主, 土层深厚, 是麻竹生长的最适区。植被以芒萁、五节芒、桃金娘和蕨类为主。
试验林于1995年春移栽母竹造林, 面积为2 hm2, 按株行距5 m×5 m设计, 密度为400株·hm-2 。竹林前期(造林后1~3 a)每年春施肥、除草1次, 4~5 a年每年春除草1次, 施肥2次, 分别于春季和8月初进行。现有竹林每丛留5株, 分别为1 a生2株, 2 a生2株, 3 a生1株。
2 研究方法 2.1 测定时间、样品确定及保护(郑郁善等, 1998; 徐志防等, 1999; 傅天明等, 1990)麻竹6月初开始发笋, 定为发笋前期, 7月底至8月中旬大量发笋定为发笋盛期, 9月上旬开始留竹, 并且发笋量较少, 定为后期。测定时间前期为6月上旬(4~10日), 盛期为8月上旬(2~10日), 末期为9月中旬(10~18日)。
测定竹株按竹林平均胸径不超过5%范围选择3 a生、2 a生、1 a生麻竹, 且3个年度竹子生长在同一丛; 分析样品全部选取阳面枝条, 并剪取其顶梢下部第2轮竹叶(2000年新长竹叶), 上、中、下3个部位按竹冠长度划分。发笋前期, 由于竹林经过冬季落叶, 刚刚重新开始郁闭, 竹冠各部位受光差异不大, 所以, 只选取中部竹叶测定。
每天采集枝条, 用湿棉花包好枝条基部, 放于保温箱中(以塑料袋包好冰块, 经测定温度在18℃左右), 并喷水保湿, 带回光合室当天分析。
2.2 叶面积测定采用美国产的Li-3000叶面积仪测定叶面积。
2.3 净光合速率和暗呼吸速率测定采用北京均方理化研究所生产的GXH-305便携式CO2 分析仪测定15000 Lux离体枝条的净光合速率; 用黑布遮光测定暗呼吸速率。
2.4 光补偿点测定测定0-20000 Lux光强下的光合强度, 测定扣除叶室吸收后到达叶面的实际光强, 以实际接受的光强计算离体枝条的光补偿点。
2.5 光呼吸速率测定通入的气体用碱石灰把CO2 吸收, 测定叶片15000 Lux光照下放出的CO2 浓度, 计算光呼吸速率。
2.6 CO2 补偿点测定采用首尾相接的闭路式气路系统, 在15000 Lux光照下待气路中CO2 浓度下降到恒定水平时即为CO2 补偿点。
2.7 光合及呼吸作用条件控制(1) 光强:用碘钨灯为光源, 交流电子稳压器稳定电源, 照度计测光强。(2)温度:用空调调节室内温度, 叶室上方用隔热水槽降温, 用温度计测定温度。(3)湿度:在叶室中用湿棉花调节湿度。(4)气源:采集离地面5 m的稳定空气。
3 结果与分析 3.1 净光合速率净光合能力反映麻竹利用太阳能的能力, 它是衡量光合能力的重要指标。从不同发笋时期来看, 各年度麻竹净光合速率最强时期在发笋盛期, 其平均值达1.9235 mgCO2 ·dm-2 h-1, 是发笋末期各年度麻竹平均值的1.21倍, 这主要是由麻竹生理周期决定。在发笋盛期, 麻竹生理机能较旺盛, 酶的活性也相对于其它时期高, 所以具有较强的光合能力。但在发笋初期, 净光合速率值出现负值, 这可能一方面是由于麻竹主要分布在水热条件较好的区域, 忍受低温能力较差, 而2000-01的持续极端低温对麻竹产生冻害, 导致麻竹的生理机制尚没有完全恢复正常; 另一方面, 由于测定净光合速率时光照强度较弱, 只有15000 Lux (因为6月份竹叶组织较幼嫩, 光照在35000 Lux时, 竹叶易卷曲, 影响测定结果), 随着光照强度的增强, 净光合速率增大, 光照强度大于25000 Lux时, 净光合速率普遍为正值。
|
|
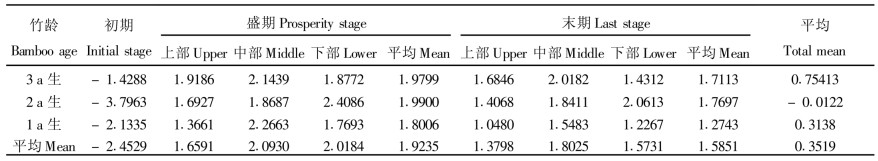
净光合能力反映麻竹利用太阳能的能力, 它是衡量光合能力的重要指标。从不同发笋时期来看, 各年度麻竹净光合速率最强时期在发笋盛期, 其平均值达1.9235 mgCO2 ·dm-2 h-1, 是发笋末期各年度麻竹平均值的1.21倍, 这主要是由麻竹生理周期决定。在发笋盛期, 麻竹生理机能较旺盛, 酶的活性也相对于其它时期高, 所以具有较强的光合能力。但在发笋初期, 净光合速率值出现负值, 这可能一方面是由于麻竹主要分布在水热条件较好的区域, 忍受低温能力较差, 而2000-01的持续极端低温对麻竹产生冻害, 导致麻竹的生理机制尚没有完全恢复正常; 另一方面, 由于测定净光合速率时光照强度较弱, 只有15000 Lux (因为6月份竹叶组织较幼嫩, 光照在35000 Lux时, 竹叶易卷曲, 影响测定结果), 随着光照强度的增强, 净光合速率增大, 光照强度大于25000 Lux时, 净光合速率普遍为正值。
不同竹龄麻竹净光合能力有一定的差异。从发笋3个时期总平均值看, 由于发笋初期影响较大, 2 a生和1 a生净光合速率总平均值都出现负值, 其中, 2 a生的麻竹达到-1.3455 mgCO2 ·dm-2 h-1, 若仅从盛期和末期看, 2 a生麻竹净光合能力比3 a生和1 a生的强, 其次为3 a生麻竹。
从盛期和末期竹冠不同部位净光合速率看, 3 a生和1 a生麻竹都呈现中间部位净光合速率值大于上部和下部竹叶, 而2 a生麻竹净光合值有随竹冠的升高而增大的趋势。但从各年生麻竹不同部位平均值看, 麻竹中部竹叶光合能力较强, 下部其次, 上部最弱。不同年度麻竹净光合速率与竹叶在竹冠中的位置关系, 是否与麻竹水分和无机养分输送及竹叶在林分中受光程度或竹叶结构有关, 目前尚不清楚, 有待于以后进一步研究。
方差分析结果表明(郎奎健等, 1989), 不同发笋时期净光合速率达显著差异(F=31.80>F0.05 (2, 4) =6.94), 而不同年度麻竹净光合速率没有达到显著差异水平(F=0.79 < F0.05 (2, 4) =6.94)。
3.2 光呼吸植物的绿色细胞在光合作用的同时, 也进行呼吸作用, 由于这种呼吸作用是在光照下才能进行, 与光合作用密切联系, 所以称这种呼吸为光呼吸。目前学者普遍认为光呼吸只是一个消费过程, 光呼吸越小, 越有利于有机物积累。从表 2看, 在麻竹不同发笋时期, 光呼吸差异较大。在发笋盛期, 各年生麻竹光呼吸最小, 只有0.8108 mgCO2 ·dm-2 h-1, 不足发笋初期的37.97%和47.15%;而在发笋初期, 光呼吸达2.1355 mgCO2 ·dm-2 h-1, 远大于其它各个时期, 这可能是由于麻竹刚受极端低温影响, 竹叶的器官和组织尚在恢复, 尚没有完全恢复正常, 而麻竹发笋末期光呼吸增大, 这可能是麻竹本身的生理特性所致。从光呼吸在不同发笋期的差异来看, 麻竹光呼吸可能不仅仅只是一个消费过程, 它一方面与植物体本身生理活动有关系, 另一方面还可能与外界环境变化密切联系, 特别是与植物抗逆性或植器官和组织的受伤后的修复有关, 但具体机理尚不清楚, 有待于进一步研究。
|
|

不同竹龄麻竹的光呼吸也具有一定差异。从表 2可看出, 在发笋盛期, 光呼吸有随竹龄的增大而增大的趋势, 但差异不大, 极值为0.1436 mgCO2 ·dm-2 h-1。在发笋末期, 2a生麻竹光呼吸速率最小, 但其值也是盛期的1.45倍; 1a生麻竹和3a生麻竹光呼速率吸相近, 为1.9895和1.9762 mgCO2 ·dm-2 h-1, 是盛期的近2.75倍和2.12倍; 在发笋初期, 2 a生麻竹光呼吸速率最大, 达2.5651 mgCO2 ·dm-2 h-1, 而1 a生和3 a生麻竹分别为1.9364和1.9050 mgCO2 ·dm-2 h-1, 这可能与麻竹受低温的影响有关。
不同竹冠部位竹叶光呼吸也具有一定差异, 呈现中部竹叶光呼吸较小, 面上部其次, 下部最大的变化趋势。
但方差分析表明, 各发笋期和各年度麻竹光呼吸速率达显著水平(F时期=7.86>F0.05 (2, 4) =6.94), 不同年度麻竹光呼吸速率差异不大(F年度=0.02 < F0.05 (2, 4) =6.94)。
3.3 暗呼吸速率麻竹的暗呼吸速率在发笋期是呈现动态变化, 不同发笋期麻竹暗呼吸速率具有较大的差异。在各个时期中, 各年度麻竹都以盛期麻竹呼吸速率为最小, 平均速率为1.2624 mgCO2 ·dm-2 h-1, 初期速率最大, 为盛期的1.81倍, 是末期的1.36倍, 明显大于其它各个时期, 这可能也与极端低温有关。
不同年度麻竹暗呼吸速率不同。从表 3可看出, 暗呼吸速率在整个发笋期的总平均值有随竹龄的增加而减小的变化趋势, 但在不同的发笋时期又有一定的变化。在初期和末期, 呼吸速率随竹龄的增加而减小, 而在盛期以2 a生竹为最小。
|
|
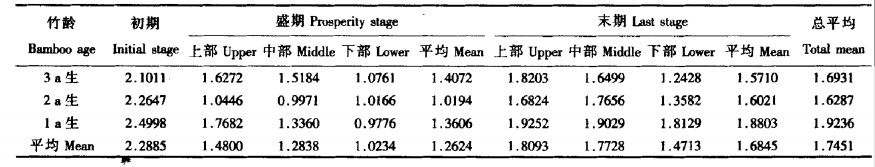
方差分析表明, 各发笋期和各年度麻竹暗呼吸速率达显著水平(F时期=33.91>F0.05 (2, 4) =6.94), 不同年度麻竹暗呼吸速率差异不大(F年度=2.84 < F0.05 (2, 4) =6.94)。
|
|
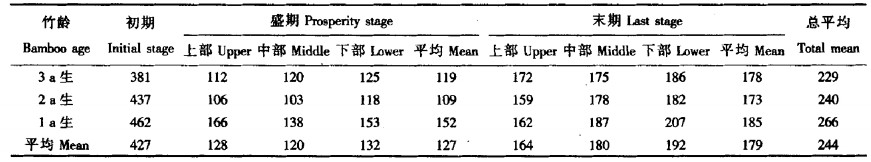
CO2 补偿点是反映植物利用低浓度CO2 进行光合作用能力的一个重要指标, 补偿点越低, 利用低浓度CO2 进行光合作用能力越强。从麻竹发笋期看, CO2 补偿点呈高—低—高变化趋势。发笋初期CO2 补偿点含量最高, 3个年度麻竹平均值达427 mg·kg-1, 而盛期最低, 只有127 mg·kg-1, 为初期的30.46%, 末期的70.95%。从CO2 补偿点也可说明麻竹在发笋盛期, 竹叶生理机能旺盛, 具有较强的利用低浓度CO2 的能力。
不同年度麻竹CO2 补偿点也具有一定的差异。尤为明显的是1 a生麻竹, 其各时期CO2 补偿点都比其它2个年度麻竹高, 这可能是由于1 a生麻竹组织较幼嫩, 在低浓度CO2 下光合能力较差; 2 a生麻竹在发笋初期CO2 补偿点比3 a生麻竹高, 但盛期和末期CO2 补偿点都相对比3 a生麻竹低, 这是否与极端低温有关, 有待进一步研究。但从方差分析结果看, 不同发笋时期和各年度麻竹CO2 补偿点达显著水平(F时期=154.11 < F0.05 (2, 4) =6.94), 不同年度麻竹CO2 补偿点差异不大(F时期=2.52 < F0.05 (2, 4) =6.94)。
从盛期和末期不同部位竹叶CO2 补偿点看, 在盛期, 2 a生和1 a生麻竹补偿点以中部竹叶较低, 3 a生麻竹随竹叶的升高而下降; 而末期各年度麻竹CO2 补偿点从上至下有逐渐升高的变化趋势。
3.5 光补偿点光补偿点反映的是植物利用弱光的能力。麻竹不同笋期光补偿点具有很大的差异。在初期, 各年度麻竹光补偿点达23436 Lux, 是盛期和末期的36.33和38.74倍, 远远高于盛期和末期, 表现较为异常, 这可能是由于极端低温, 影响麻竹光合作用。在其它2个时期, 3 a生和2 a生麻竹光补偿点呈现出末期大于盛期, 但1 a生麻竹相反。从各时期3个年度麻竹光补偿点平均值看, 从初期到末期呈逐渐降低趋势, 末期最小, 平均为605 mg·kg-1, 盛期次之, 初期为最大。
|
|
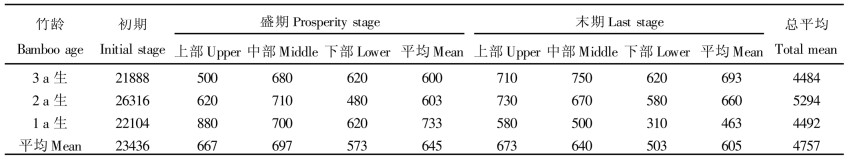
不同年度麻竹光补偿点具有一定的差异。从3个发笋时期总平均值看, 2 a生麻竹光补偿点较高, 而3 a生和1 a生麻竹较低。但不同年度麻竹光补偿点在各个时期有一定的波动, 初期以2a生麻竹为最高, 而盛期光补偿点有随竹龄的增大而减小的趋势, 末期相反。
从不同部位麻竹叶片光补偿点看, 总体上有随部位的升高而减小的趋势。从各年度表现上, 1 a生麻竹光补偿点从上至下逐渐下降, 且盛期明显高于末期; 但在3 a生和2 a生麻竹中, 盛期以中部竹叶补偿点为较高, 而末期也呈现出随竹叶着生高度的下降而明显下降的趋势。
方差分析结果表明, 不同发笋时期光补偿点达显著水平(F时期=250.08>F0.05 (2, 4) =6.94), 不同年度麻竹光补偿点差异不大(F时期=1.01 < F0.05 (2, 4) =6.94)。
3.6 光合系数光合系数表示在最适宜条件下, 从全部光合作用收入中, 叶器官为本身呼吸所必需消耗的量。由于总光合速率不能直接测定, 常用净光合速率与暗呼吸之和求出近似值。它表明光合速率与呼吸作用配合情况, 当高光合速率与较低呼吸配合时, 光合作用产额高, 光合系数就大。用光合系数KF表示为:
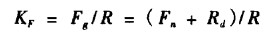
|
其中:Fg、R、Fn、Rd分别表示总光合速率、总呼吸速率(光呼吸速率和暗呼吸速率之和)、净光合速率和暗呼吸速率。
从表 6可知, 不同发笋时期光合系数不同。在发笋盛期光合系数较大, 3个年度麻竹光合系数平均值达1.5772, 而到末期减小为0.9907, 但在初期, 光合系数出现负值, 主要是而净光合为负值, 特别是2 a生麻竹光合系数出现负值, 这主要是由于净光合速率出现负值且大于暗呼吸所至。从不同竹龄麻竹看, 麻竹光合系数随竹龄的增大而增大, 这主要是由于2 a生麻竹初期光合系数为值, 若仅从盛期和末期看, 2 a生麻竹光合系数最大, 3 a生麻竹其次, 1 a生麻竹较小。但无论如何, 这说明3 a生麻竹生理机能尚处于较旺盛期, 目前有些地方麻竹笋用林经营中, 只留1、2 a生麻竹, 每年留养新竹, 从麻竹的光合生理看, 有欠妥之处。从不同部位叶片看, 盛期以中部叶片光合系数较大, 上部次之, 下部最小, 末期有随竹叶的升高而增大的变化趋势。
|
|
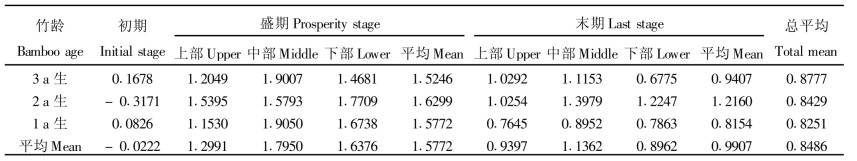
方差分析表明, 各发笋期和各年度麻竹光合系数达显著水平(F时期=35.83>F0.05 (2, 4) =6.94), 不同年度麻竹光呼吸速率差异不大(F时期=0.04 < F0.05 (2, 4) =6.94)。
4 小结与讨论麻竹在发笋盛期生理机能比发笋初期和末期旺盛, 具有较强的同化能力, 其净光合速率较大, 光呼吸、暗呼吸、CO2 补偿点和光补偿点低, 光合系数也较大。不同年度麻竹光合和呼吸特性由于发笋初期表现异常, 具有一定的波动, 盛期和末期上2 a生麻竹具有较强的光合能力和较低的呼吸, 光合系数明显高于其它年度麻, 3 a生次之, 1 a生麻竹光合能力较弱。
麻竹不同部位竹叶光合特性不同, 净光合速率以中部竹叶为最大, 下部其次, 上部最低; 光呼吸变化波动较不明显; 暗呼吸速率从上到下有减小的趋势; CO2 补偿点在盛期以中部较低, 3 a生麻竹从上至下有升高的趋势, 末期各年度麻竹都随竹叶的升高而下降; 光补偿点从上至下有降低的趋势; 光合系数在盛期以中部较大, 末期随竹冠部位的升高而增大。
麻竹枝条上的叶1年4季在不断的生长, 所以从顶梢至下部竹叶成熟度不一致, 其光合速率也可能不同, 另外, 由于竹叶在强光照下容易卷曲, 为了使试验结果具有可比性, 试验采用15000 Lux较低光强, 选取向阳面枝条顶梢下第2轮竹叶为试验材料进行比较其光合和呼吸速率, 试验结果只能代表某一时刻同一部位竹叶的光合和呼吸状况, 与自然界一日中不断变化的光强, 不同位置竹叶光合和呼吸速率有一定差异。
从麻竹的光合和呼吸特性测定结果看, 麻竹在发笋盛期生理机能旺盛, 在前期应做好水肥的管理, 以保证发笋期养分的供应, 增加竹笋产量。从各年度麻竹生理特性看, 2、3a生麻竹光合能力较强, 因此, 在郁闭笋用麻竹林经营过程中, 是否采取目前一些多地方每年留竹, 且只留1、2a生麻竹的经营方法有待进一步改进。因为麻竹笋用林经营以笋为主要产品, 而每年的留竹, 消耗大量的养分, 也缩短了发笋期, 这样可能会降低竹林发笋量, 竹林经济效益降低。
傅天明, 等. 1990. 夏大豆生育期间光合特性的研究. 中国农业科学, 23(6). |
黄克福编.竹林培育技术.福州: 福建科学技术出版社, 1992
|
郎奎健, 唐守正. 1989. IBMPC系列程序集. 北京: 中国林业出版社, 102-103.
|
梁鸿木, 陈学魁. 1998. 麻竹单株生物量模型研究. 福建林学院学报, 18(3): 260-262. DOI:10.3969/j.issn.1001-389X.1998.03.017 |
肖贤坦, 马占兴, 朱培琨. 1996. 麻竹丰产林培育技术与应用. 竹类研究, (1): 57-60. |
徐志防, 罗广华, 王爱国, 等. 1999. 光合作用的光抑制与光合器官的活性代谢. 植物学通讯, 35(4): 325-332. |
郑郁善, 洪伟, 邱尔发, 等. 1998. 营养液对毛竹叶片效应的研究.Ⅱ.营养液对叶片光合性状影响. 林业科学, 34(专刊1): 95-99. |
周本智, 等. 1999. 闽南麻竹人工林地上部分现存生物量的研究. 林业科学研究, 12(1): 47-52. DOI:10.3321/j.issn:1001-1498.1999.01.009 |
周本智, 等. 1999. 麻竹出笋和高生长规律的研究. 林业科学研究, 12(5): 461-466. DOI:10.3321/j.issn:1001-1498.1999.05.003 |