 2001, Vol. 37
2001, Vol. 37文章信息
- 谢锦升, 林瑞余, 黄荣珍, 陈银秀, 杨玉盛.
- Xie Jinsheng, Lin Ruiyu, Huang Rongzhen, Chen Yinxiu, Yang Yusheng.
- 严重退化红壤上恢复的马尾松—灌木混交林群落能量格局的研究
- STUDY ON ENERGY PATTERN OF RECOVERING THE P. MASSONIANA AND SHRUBS MIXED FOREST ON THE SERIOUS DEGRADATION RED SOIL
- 林业科学, 2001, 37(专刊1): 131-136.
- Scientia Silvae Sinicae, 2001, 37(专刊1): 131-136.
-
文章历史
- 收稿日期:2001-01-08
-
作者相关文章


长汀河田镇是我国南方花岗岩地区最典型的水土流失区之一, 严重的水土流失使山地土层母质裸露, 土壤肥力下降, 植被生长不良, 生态环境恶化, 严重阻碍了该地区的可持续发展和人民生活水平的提高。为了治理严重的水土流失, 重建功能良好的生态系统, 从20世纪80年代起各级主管部门相继采取许多治理措施, 使河田水土流失得到初步控制, 以小流域为单元的一些退化生态系统得到一定程度的恢复。杨玉盛等(1996;1998;1999)已对严重退化土地不同治理措施采取后的土壤抗蚀性、土壤物理化学性质及生物多样性等方面进行了研究, 本文对严重退化土地上恢复的马尾松—灌木混交林群落的的能量生产能力进行探讨, 这对类似地区开展生态恢复实践具有一定指导意义。
1 试验地概况福建河田属中亚热带季风气候, 年均气温19℃, 7月份极端最高气温39.8℃, 地表极端最高温达76.6℃ (1983-07-23), 年均降雨量1628.2 mm, 其中4~6月降雨量占全年的52.2%, 且降雨强度大。土壤为粗晶花岗岩风化发育的山地丘陵红壤, 含沙量大(> 1 mm石砾占45%左右), 风化层深厚。该镇水土流失面积占44.65%, 其中强度流失面积占流失总面积的58.93% (1983年调查)。
马尾松—灌木混交林:试验地点位于八十里河, 1981年冬在原侵蚀地采用小水平沟整地, 沟距1.5 m, 面宽0.6 m, 深0.4 m, 底宽0.4 m。整地时保留原有马尾松、木荷(十分稀少)等乔木, 沟长2~3 m留一土埂, 每hm2实际沟长约23.3 m。每m沟长施基肥棉籽饼0.1 kg, 钙镁磷肥0.2 kg, 猪粪1 kg; 1982年春季在马尾松林下种胡枝子和紫穗槐, 种植当年5、8月及第2年5月追肥3次(尿素、棉籽饼等)。每年砍伐胡枝子和紫穗槐覆盖林地。标准地坡向NE35°, 坡度14°, 马尾松平均树高10.95 m, 平均胸径9.0 cm, 密度3425株·hm-2, 林分郁闭度0.90, 林下植被主要为胡枝子和芒萁, 亦出现马唐、苔藓、地衣等植物, 林下植被层盖度0.30, 苔藓层盖度0.70。土壤A+AB层大约1~2 cm。
对照:以邻近严重侵蚀地为对照, 坡向NE30°, 坡度12°, 对照地内土壤侵蚀严重, 浅沟发育, 地表植物以马尾松小老头树和少量芒萁和野古草为主, 马尾松密度925株·hm-2, 平均树高0.8 m, 平均地径3.1 cm, 树龄20 a左右, 林分郁闭度0.2, 植被盖度0.1, 无植被地方近于光板地, 由于土壤侵蚀严重, 立地条件差, 部分马尾松根系裸露, 表层土壤有机质0.5~4.3 g·kg-1, 全N 0.04~0.16 g·kg-1。
2 研究方法于1999~2000年分别在马尾松—灌木混交林和对照地内设置20 m×20 m标准地3块, 在每块标准地进行每木检尺, 并以平均胸径选1株标准木, 按Monsi分层切割法测定乔木层地上部分生物量, 采用全挖法测定根系生物量, 乔木层各器官生物量净增量采用相对生长法进行推算。灌木和草本层生物量测定采用小样方法(5个, 1 m×1 m), 调查时灌木按枝、叶和根, 草本按地上和根分别称量, 取样测定含水率。枯枝落叶层数量测定亦采用小样方法, 与林下植被层调查同步进行(冯宗炜等, 1999; 杨玉盛等, 1998)。苔藓测定地面生苔藓的生物量, 取5个面积为20 cm×20 cm的全部苔藓植物, 分别置于塑料袋内直接称重, 换算的每m2鲜重, 将样品放于通风室温下干燥1周以上, 称重换算得干重(曹同等, 1995)。其生物量净生长量的测定采用灌木和草本层的生物量现存量分别除以其平均年龄而得, 本文中灌叶、草茎叶、草根、苔藓的平均年龄按2 a计, 灌枝、灌根的平均年龄按8 a计(林益明等, 1996; 杨玉盛等, 1998)。凋落物测定通过设置框架定期收集。各植株样品经烘干、磨碎, 用HWR-15恒温式微机量热计测定其干重热值, 重复2~3次。
3 结果与分析 3.1 马尾松—灌木混交林群落各组分的干重热值含量植物热值是指单位重量干物质在完全燃烧后所释放出来的热量值, 群落各组分样品的热值测定是群落能量生产研究的基础(林益明等, 1996)。由表 1可见, 各群落乔木层地上部分各组分的干重热值(GCV, Gross Caloric Value)的大小顺序均为叶 > 枝 > 干 > 皮, 这与前人的研究结果较相似, 表明叶片的热值含量高。Howard-Williams (1974)认为叶的高热值现象与在贫瘠的土壤条件下叶片高能化合物如粗脂肪和蛋白质等的积累有关。河田的土壤非常贫瘠, 马尾松树叶热值亦最高, 这可能是在贫养条件下对环境适应的结果。混交林地上部分各器官干重热值含量均分别大于对照相应器官的, 这可能与各器官中高能有机物含量比对照的高有关。乔木层地下部分 > 0.2 cm根的干重热值高于 < 0.2 cm细根, 而在 > 0.2 cm根中, 径级较大的根干重热值含量有大于较小径级的趋势。对照的根系干重热值一般比马尾松—灌木混交林的大, 这可能与对照地马尾松个体小、林分郁闭度低、根系出露表土层(冲刷所致)有关。
|
|
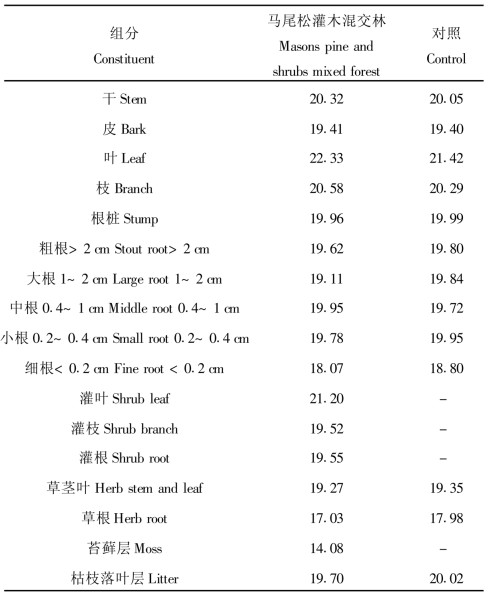
从地上部分与地下部分相比较可看出, 群落乔木层地上部分热值含量则均高于地下部分, 这与地上部分和地下部分所处的环境和所担负的功能有关。
混交林灌木层GCV大小为灌叶 > 灌根 > 灌枝, 而混交林和对照群落草本层的GCV大小为草茎叶 > 草根, 且对照的茎叶和根均比混交林的草本层相应器官的高, 这与对照林分郁闭度低有关。混交林群落各层次的GCV大小则为乔木层 > 灌木层 > 草本层 > 苔藓层, 这与各层次植物所处生境的光照强度有关, 从乔木层、灌木层、草本层到苔藓层光照强度减弱, 因而干重热值下降。这也说明了群落中不同层次植物的生存策略差异, 在群落中所处的层次越低, 由于光照强度越弱, 植物所能利用的光能减少, 因而植物必须尽量减小生产单位干重物质所耗费的能量, 这种结果与落叶松人工林和甜槠林群落相似(林益明等, 1996; 刘世荣等, 1992)。
混交林枯枝落叶层的干重热值低于对照的, 两者干重热值均低于相应乔木层的枝、叶, 可能与枯枝落叶遭微生物分解后高能物质流失而纤维素数量比例增加有关。而乔灌混交林群落枯枝落叶热值低于对照的, 这可能与其林内环境有利于其枯枝落叶分解有关。治理后土壤微生物数量是对照的23.7倍(杨玉盛等, 1998), 表明乔灌混交林有利于促进土壤微生物活动, 从而加速了枯枝落叶的分解。
3.2 马尾松—灌木混交林群落能量现存量群落的能量现存量是指某一给定时刻、单位面积上, 群落中含能产品中所贮藏的能量(林益明等, 1996)。马尾松—灌木混交林群落的能量现存量达25.89×104 kJ·m-2, 是对照群落的156.1倍(表 2)。乔灌木混交林群落乔木层的能量现存量占总量的98.1%, 是对照的259.9倍; 干皮和枝叶的能量现存量占乔木层总量分别为59.6%和26.1%, 而对照的则分别为29.9%和56.9%, 表明与对照的相比, 混交林乔木层投入更高比例的能量用于构建结构组织(干皮), 而对照的则相反, 由于群落光合作用能力极低, 获取的能量首先用于构建光合作用器官(叶、枝), 以提高群落的光合作用能力。混交林灌木层、草本层和苔藓层的能量现存量仅占总量的0.3%和0.5%和1.1%;而对照的由于立地条件极差, 虽然林分郁闭度很低, 但是林下植被中未出现灌木层和苔藓层, 仅出现少量野古草、芒萁等草本, 其能量现存量亦仅为乔灌木混交林的48.7%, 但由于其乔木层光能利用率极低, 使其草本层在群落能量中所占比例达39.9%, 远比混交林草本层所占的比例为高。乔灌混交林地表苔藓层的出现, 一方面利用林内透射光或散射光构建其生物量; 另一方面可有效地防止光线直接照林地, 从而大大缓和林内土壤温度的变化, 这对土壤生态系统功能的恢复有十分重要的意义。
|
|
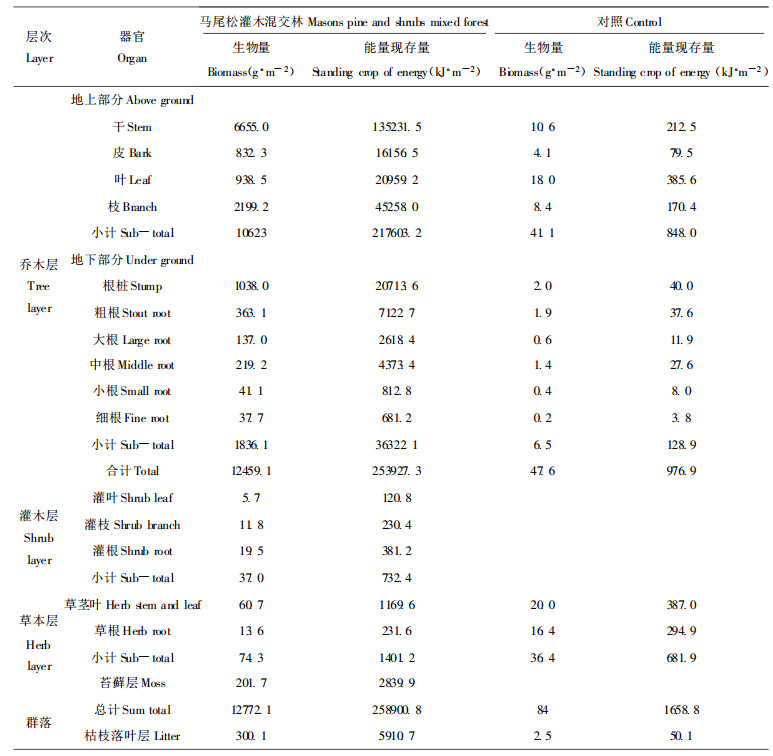
枯枝落叶层能量的多少, 标志着群落对土壤亚生态系统输入能量的多寡; 同时, 枯枝落叶层可滞蓄降水, 遮挡日光直射, 对提高土壤的含水量, 为土壤中的生物体创造适宜的生存环境等有积极意义。混交林和对照的枯枝落叶层分别占群落总能量的2.2%和2.9%, 但混交林枯枝落叶层的总能量则是对照的118.0倍, 表明对照地以枯枝落叶形式向土壤输入的能量(有效能)极少, 而更多地以光辐射能(无效能)直接进入土壤, 这对土壤生态系统的恢复与重建十分不利。
乔灌混交林群落的层次比对照的仅由乔木层和草本层组成多了灌木和苔藓层, 因而可多层次地截留光能, 增加有机质的合成, 这对促进土壤的生态重建是有重要作用。而对照地群落层次单调, 地表植被覆盖度和群落叶面积指数极低, 对太阳能截获能力差, 大部分太阳辐射为地表土壤所吸收, 使得白天地表温度过高(最高时可高达76.6℃), 昼夜温差过大, 地表水分蒸发强烈, 土壤过度干旱, 植物难以生长, 生态系统处于严重退化过程中。因此, 降低太阳光向地表土壤输入过多的能量(无效能), 则成为中亚热带严重退化地生态重建的关键。而增加地表植被的覆盖, 特别是构建多层次结构, 提高光能利用率和群落的生物量, 不但可使群落对能量环境的调节能力不断增强, 从而为退化系统的生态重建提供一个较为适宜和稳定的热辐射环境, 而且可增加光合有机质的积累和对土壤亚生态系统的有机物(有效能)的输入, 从而促进退化生态系统的恢复。
3.3 群落的能量年存留量、归还量与净固定量群落的能量年存留量是群落中各组分的生物量年净增量与相应干重热值乘积的总和(林益明等, 1996)。乔灌混交林群落年能量存留量达27092.03 kJ·m-2a-1, 是对照群落的59.6倍, 而乔木层能量年存留量是对照的218.6倍(表 3), 但均低于两者生物量现存量的比值, 说明治理措施实施近20 a左右, 由于对立地条件改良能力有限, 相关措施一旦停止使用, 处理的马尾松生长将呈下降趋势(未刊资料)。混交林和对照草本层的能量年存留量占群落总能量存留量的分别为2.59%和75.00%, 但数量都极小, 混交林的是对照的2.1倍; 说明混交林由于高度郁闭, 限制林内草本的生长, 而对照地则干旱、贫瘠太突出, 使得其灌木难以生长, 就是草本种类和数量亦是极低的。
|
|

群落的能量年归还量是指群落年凋落物量与其平均干重热值乘积(林益明等, 1996)。混交林群落的能量年归还量为5825.01 kJ·m-2a-1, 是对照(45.26 kJ·m-2a-1)的128.7倍, 可见混交林以凋落物等有机物形式向土壤亚生态系统归还能量远高于对照的, 每年大量含能的有机物归还给土壤, 这对退化生态系统土壤有机质的恢复, 促进土壤微生物和小动物活动, 土壤肥力恢复等具有重要意义。
群落能量净固定量是指单位时间、单位面积的某一群落通过光合作用所固定的总能量扣除群落因自身呼吸所消耗能量后所剩余的能量。因而本文中群落的能量净固定量包括群落年能量存留量和年能量归还量。乔灌木混交林群落的年能量净固定量达32917.04 kJ·m-2a-1, 是对照(499.82 kJ·m-2a-1)的65.9倍。在混交林群落的年能量净固定量中, 用于群落自身增长的能量(能量年存留量)占82.3%, 而释放到土壤亚生态系统的能量(能量年归还量)则占17.7%。与其它一些陆生群落相比, 乔灌木混交林群落的年能量净固定量高于甜槠林(26856.2 kJ·m-2a-1) (林益明等, 1996)、雨绿林(26334 kJ·m-2a-1) (Helmut, 1975)、落叶松人工林(26435.2 kJ·m-2a-1) (刘世荣等, 1992), 但略低于热带雨林(34276 kJ·m-2a-1) (Helmut, 1975)和中亚热带的杉观混交林(35996 kJ·m-2a-1, 未刊资料), 而与中亚热带的杉木纯林(32429 kJ·m-2a-1, 未刊资料)接近, 表明乔灌木混交林群落具有较高的能量生产力。
一般用群落每年净固定能量(NEP)占当年太阳光合有效辐射能(PhAR)来推算群落太阳能转化效率(ECE), 即ECE%=NEP%/PhAR×100% (林益明等, 1996)。据当地气象站测定, 该地的多年太阳辐射量(SRE)为4531000 kJ·m-2a-1, 则太阳的光合有效辐射(PhAR= SRE×0.47)为2129570 kJ·m-2a-1。经计算, 乔灌木混交林群落的太阳能转化率为1.55%, 低于红树林群落(2.01%~3.01%) (林光辉等, 1988)、落叶松人工林(2.30%) (刘世荣等, 1992)和热带人工林(2.35%~5.07%) (邓瑞文等, 1985), 但高于中亚热带的甜槠林(1.30%) (林益明等, 1996)及栎林(0.73%) (Rerners, 1971), 而与中亚热带杉观混交林(1.60%)和杉木纯林(1.44%)的接近, 表明乔灌木混交林群落具有较高的太阳能转化率, 同时也表明采取乔灌混交进行治理, 群落的能量生产能力得到恢复, 治理的效果较好。而对照群落的太阳能转化率仅为0.023%, 可见对照群落的太阳能转化率极低, 说明对照生态系统是极度退化的, 但同时也说明对照具有较大的光能利用潜力, 只要经过合理治理, 可以大幅度提高群落的生产能力。
4 小结乔灌木混交林乔木层地上部分各器官干重热值均大于相应对照的, 且以叶的最高, 而地下部分干重热值则低于对照的。乔灌木混交林群落各层次的GCV大小为乔木层 > 灌木层 > 草本层 > 苔藓层。
乔灌木混交林群落的能量现存量达258900.8 kJ·m-2, 是对照群落的156.1倍。乔灌木混交林群落由乔木层、灌木层、草本层和苔藓层组成, 各层次能量现存量分别占群落的98.1%、0.3%和0.5%和1.1%。而对照群落仅由乔木层和草本层组成, 乔木层和草本层分别占群落的60.1%和39.9%;由于对照群落植被盖度极小, 光能转化效率很小(0.023%), 群落内昼夜温差大、土壤干旱, 生态系统处于严重退化过程中。而增加植被覆盖, 则是中亚热带严重退化系统生态重建的关键。
乔灌木混交林群落年能量存留量达27092.03 kJ·m-2a-1, 是对照群落的59.6倍; 能量年归还量为5825.01 kJ·m-2a-1, 是对照的128.7倍, 年能量净固定量达32917.04 kJ·m-2a-1, 是对照的65.9倍, 其群落的太阳能转化率为1.55%, 表明采取乔灌混交治理近20 a后, 群落的能量生产能力得到恢复, 治理的效果较好。
曹同, 高谦, 傅星, 等. 1995. 长白山森林生态系统中苔藓植物的生物量. 生态学报, 15(增B): 68-73. |
邓瑞文, 陈天杏, 冯咏梅. 1985. 热带人工林的光能利用与生产量的研究. 生态学报, 5(3): 231-240. |
冯宗炜, 王效科, 吴刚. 1999. 中国森林生态系统的生物量和生产力. 北京: 科学出版社.
|
林光辉, 林鹏. 1988. 海莲、秋茄两种红树群落能量的研究. 植物生态学与地植物学学报, 12(1): 31-39. |
林益明, 林鹏, 李振基, 等. 1996. 福建武夷山甜槠群落能量的研究. 植物学报, 38(12): 989-994. |
刘世荣, 王文章, 王明启. 1992. 落叶松人工林生态系统净初级生产力形成过程中的能量特征. 植物生态学与地植物学学报, 16(3): 209-219. |
杨玉盛, 何宗明, 林光耀, 等. 1996. 不同治理模式对严重退化红壤抗蚀性影响的研究. 土壤侵蚀与水土保持学报, 2(2): 36-42. |
杨玉盛, 何宗明, 林光耀, 等. 1998. 退化红壤不同治理模式对土壤肥力的影响. 土壤学报, 35(2): 276-282. DOI:10.3321/j.issn:0564-3929.1998.02.017 |
杨玉盛, 何宗明, 邱仁辉, 等. 1999. 红壤严重退化生态系统不同恢复和重建措施的植物多样性和地力恢复的研究. 生态学报, 19(4): 490-494. DOI:10.3321/j.issn:1000-0933.1999.04.010 |
杨玉盛. 1998. 杉木林可持续经营的研究. 北京: 中国林业出版社.
|
Helmut Lieth, Robert H Whittaker.生物圈的第一性生产力(王业蘧译).北京: 科学出版社, 1985
|
Howard-Williams C. 1974. Nutritional quality and caloric value of Amazonian forest litter. Amazoniana, 1: 67-75. |
Rerners W A. 1971. Sructure and energetics of three Minnessota forests. Ecol. Monogr., 42: 71-94. |