 2001, Vol. 37
2001, Vol. 37文章信息
- 江希钿, 杨锦昌, 王素萍.
- Jiang Xidian, Yang Jinchang, Wang Suping.
- 同龄纯林自然稀疏密度变化的研究
- A STUDY ON THE SELF-THINNING OF DENSITY VARIATION FOR EVEN-AGED PURE STANDS
- 林业科学, 2001, 37(专刊1): 84-89.
- Scientia Silvae Sinicae, 2001, 37(专刊1): 84-89.
-
文章历史
- 收稿日期:2001-01-08
-
作者相关文章


自然稀疏现象是同龄纯林自然生长过程的一大特征, 是植物种群内林木竞争反馈调节的结果。近年来, 许多研究者采用模型技术来描述同龄纯林的自然稀疏规律, 提出了一些描述密度变化过程的理论模型(张大勇, 1985; 唐守正, 1993; 方精云, 1995; Hozumi, 1997; 1983; 刘金福等; 1998), 取得了许多可喜的研究成果。但由于自然稀疏规律的重要性和复杂性, 它们仍然是人们讨论的焦点之一。从理论上说, 一个良好的自然稀疏模型, 不仅要有一定的理论基础, 而且要应用方便。本文在前人研究的基础上, 根据林分生长和密度理论, 提出了一个新的描述一般同龄纯林林分在自然稀疏过程中密度变化规律的数学模型, 并用实际数据进行了验证, 旨在为林分密度管理与控制提供科学的依据。
1 资料收集在闽北地区生长发育正常的杉木人工林各类型林分中设置标准地, 按常规方法测算标准地的各项调查因子, 共计378块临时标准地和75块固定标准地。固定标准地测定次数为3 ~ 5次, 间隔期为2 ~ 5 a, 初测年龄分别为9、11、13 a, 最后一次测定的年龄分别为18、19、25 a。当时设置固定标准地的目的是为了研究不同间伐强度对林分生长的效应, 故有15块为对照标准地, 未进行过抚育间伐; 非对照标准地也仅在初设时进行过一次间伐, 以后就不再间伐; 对于间伐标准地, 我们选择经过3 ~ 5 a的生长而恢复自然状态的作为本次研究的基本材料。
2 最大密度林分自疏过程的密度变化模型 2.1 模型的构建植物种群在各阶段可有不同的数量, 但生物量积累随时间的变化是和单株个体重量的积累紧密相关的, 其增长曲线为S型。对于这种S型的增长曲线, 可用Gompertz方程表描述。
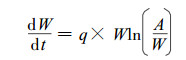
|
(1) |
式中:W为植物平均重量, A为W的上限值, t为时间, q为增长率。
方程(1)的拐点在A/e, 即最大增长速率在容纳量一半之前达到。但是最大增长速率也可能在容纳量一半之后达到。因此, 宜采用下式来描述植物种群的增长曲线。
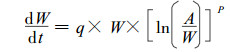
|
(2) |
P为一常数, (2)式建议植物种群的最大增长率在A/eP处达到, 即如果P >1, 那么最大增长速率在A/e之前达到, 反之在A/e之后。因此, 方程(2)实际上包括了Gompertz方程, 可以认为是植物种群生物量变化的一个较好的数学描述式。
对(2)式取积分, 并按生长方程应满足t =0时W =0的初始条件, 即生长曲线通过原点这一特性, 可得:

|
(3) |
此即Korf生长方程(李凤日, 1997), 其中: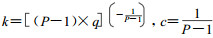
植物种群自然稀疏现象的主要特点是:通过对植物种群的生长、死亡等过程的自我调节和控制, 使种群的数量维持在生态环境可容纳的最大水平上。对于最大密度的林分, 植物种群的平均个体重量W与密度N存在幂函数关系(Yoda等, 1963), 即:
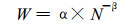
|
(4) |
式中:α为不随生长阶段和立地条件而改变的常数, β为参变量。(4)式又被称为最大密度线, 在最大密度线上, 要描述植物种群在自疏过程中的密度随时间的变化规律, 只需将方程(4)和(3)联立, 即:
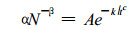
|
由此得
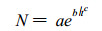
|
(5) |
其中: 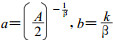
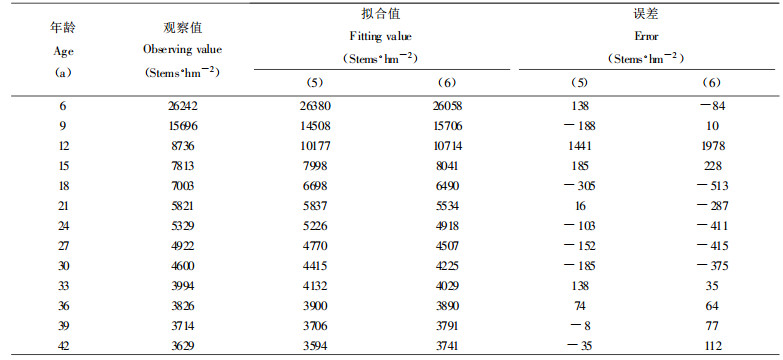
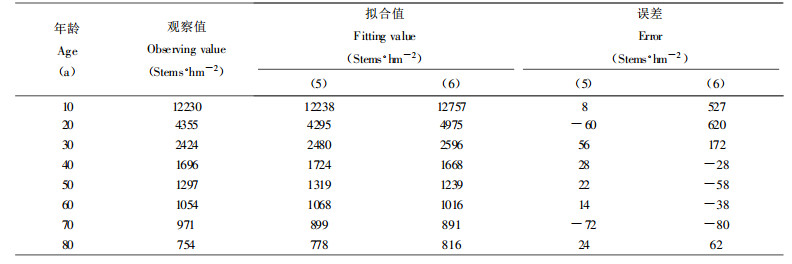
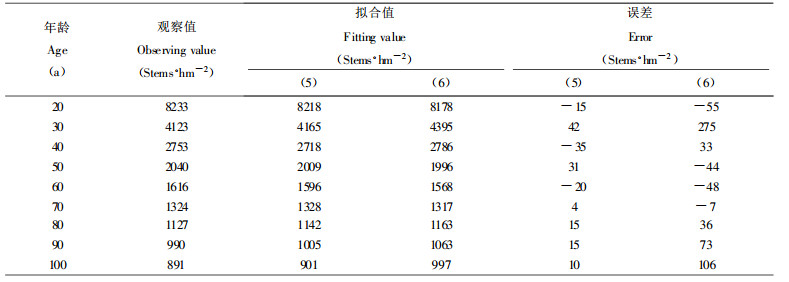
验证同龄纯林自然稀疏过程中密度变化规律的数学模型, 最好采用同一林分已连续发生大量自然稀疏时各年龄阶段所观察到的存活株数密度, 由于本次收集的杉木人工林标准地材料, 缺乏这种数据, 故引用张大勇等(1985)所调查的山杨林(Populus tremula)和云南松林(Pinus yunnanensis)自疏过程的株数观察值, 对(5)式进行验证, 各参数及相关指数R2和剩余标准差S求解结果如下:
山杨林:a =1106.227, b =7.9834, c =0.5152, R2 =0.9925, S =531株·hm-2
Ⅱ地位级云南松林:a =3.5182, b =12.8733, c =0.1983, R2 =0.9999, S =42株·hm-2
Ⅳ地位级云南松林:a =49.1509, b =14.6641, c =0.3513, R2 =0.9999, S =23.8株·hm-2
为便于比较, 将张大勇等(1985)提出的最大密度林分自然稀疏密度变化模型及参数和剩余标准差S也在此列出。

|
(6) |
(6) 式中各参数为:山杨林b1 =3529, b2 =0.1331, b3 =0.1027, b4 =16, t0 =6, S =611株·hm-2; Ⅱ地位级云南松林b1 =690, b2 =0.2, b3 =0.042, b4 =16, t0 =10, S =297株·hm-2; Ⅳ地位级云南松林:b1 =850, b2 =0.152, b3 =0.034, b4 =16, t0 =20, S =106株·hm-2。
2种模型的拟合对比如表 1、表 2和表 3所示。不难看出, 本文提出的描述最大密度林分自然稀疏的密度变化模型(5)式, 明显优于张大勇等(1985)提出的(6)式。
|
|
|
|
|
|
(5) 式描述了最大密度林分自然稀疏过程的密度变化规律, 但在现实林分中, 大量的是未达到最大密度的一般林分。因此, 研究一般林分自疏过程的密度变化规律更具有普遍性, 对指导林分密度管理有重要作用。
3.1 模型的导出在林分的自然生长过程中, 随着年龄和直径的增大株数不断减少, 对于最大密度的林分, 株数减少的相对速率为:
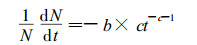
|
(7) |
(7) 式由(5)式微分而来, 描述了最大密度的林分株数减少的相对速率与林龄的关系, 对于未达到最大密度的一般林分, 各年龄阶段株数减少的相对速率还与现实林分的密度有关。因此一般林分株数随年龄增加而减少的相对速率可表达为:
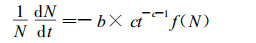
|
(8) |
其中f(N)为密度(N)的函数, 在同样年龄时, 密度越大, 自然稀疏越快, 所以f(N)是N的增函数。当N =Nmax时, 现实林分成为最大密度的林分, 这时(8)式应等于(7)式。不妨假设f(N)=(N/Nmax)θ, 它表示现实林分接近最大密度的程度。当某一年龄的最大密度由(5)式确定时, (8)式可写成:

|
(9) |
此式即为一般林分的自然稀疏方程式, 它描述了一般林分, 包括最大密度林分在自然稀疏过程中, 林分株数与年龄之间的关系, 其中θ是与树种和生境有关的常数, 可称为自稀疏指数。作为判例, 对于最大密度的林分, N =Nmax, 即N =N/aeb/tc =1, (9)式变为(7)式。解微分方程(9)式, 可得:
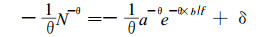
|
(10) |
其中δ是由初始条件确定的积分常数, 当t =t0, N =N0时
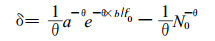
|
(11) |
将其代入(10), 得:
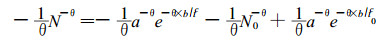
|
(12) |
整理后为:
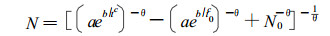
|
(13) |
(13) 式即为本文所提出的同龄纯林一般林分自然稀疏过程的密度变化模型, 它描述了某一生境条件下一般林分随年龄增加而存活株数不断减少的变化规律。
3.2 参数估计在(13)中, 需要估计的参数共有4个, 由于a、b、c 3个参数反映的是最大密度林分在自然稀疏过程中, 存活株数随年龄变化的参数。因此, a、b、c 3个参数和自稀疏指数θ应分开估计。
3.2.1 a、b、c参数的估计对于a、b、c 3个参数, 最好采用同一类型林分在自然生长过程中, 已连续发生大量自然稀疏所观察到的各年龄存活株数, 按(5)式进行估计。但是, 要获得高密度的林分在自然稀疏过程中的实际密度材料, 在实践很难做到的。我们认为, 解决的办法是采用空间序列来代替时间序列, 即选取同一生境在不同地段上不同年龄的高密度林分组成种群自然稀疏过程密度变化的时间序列。根据我们整理的闽北地区材料, 选择16 m指数级生长在不同地段上不同年龄未经人为干扰且密度尽量大的32块标准地, 组成杉木人工林种群密度变化的时间序列, 采用改进单纯形法对(5)式进行优化拟合, 得到杉木人工林最大密度林分在自然稀疏过程中密度变化模型:
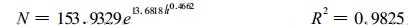
|
(14) |
该式描述了同龄纯林最大密度的林分, 单位面积株数与年龄的数量关系, 表明随着林分年龄的增加, 杉木人工林株数逐渐减小, 符合植物种群自然稀疏规律。
3.2.2 自稀疏指数θ的估计(13) 式描述的是一般林分自疏过程密度和年龄的关系。因此, 自稀疏指数θ的估计, 应采用固定标准地的两次观察资料, 若一个样地经过多次复测, 可以把相邻两次的观测值作为一对复测资料, 组成n对样本。

|
其中t0i、N0i表示第i标准地初测年龄和单位面积上的株数, ti、Ni表示第i标准地复测年龄和单位面积上的株数, 以残差平方和Q最小, 即
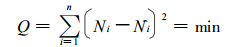
|
为目标函数, 迭代求解自稀疏指数θ值, 迭代求解的方法有多种, 一种简便实用的方法是黄金分割法, 在给定区间内, 迭代求其最优的θ值。
利用所收集的固定标准地, 选择16 m指数未经间伐或间伐后已经过3 ~ 5 a时间恢复到自然生长状态的77对复测材料, 迭代求得θ的估计值为4.9319, 这样就得到杉木人工林一般林分自然稀疏过程的密度变化模型。
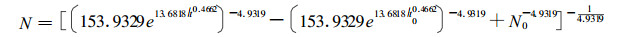
|
(15) |
为说明本文所建立的杉木人工林自然稀疏过程密度变化模型的准确性和优越性, 现应用John (1994)、Clutter(1980)、Pienaar(1981)提出的存活株数模型, 分别模拟闽北杉木人工林种群自然稀疏规律, 见表 4。由于本文所构造的杉木人工林自然稀疏模型理论基础较充分, 可以客观地反映种群自然稀疏过程的密度变化规律, 因此拟合效果明显优于其它模型。
|
|

最大密度法则和Korf生长方程的结合, 为描述同龄纯林在自然稀疏过程中的各个生长发育阶段林分所能承受的最大密度提供了理论依据, 也为编制标准收获表提供了技术手段。
从植物种群各个生长发育阶段所能容纳的最大密度出发, 推导出一般林分在自然稀疏过程中密度变化的数学模型, 有较强的机理性和较大的适用性。4种自然稀疏模型拟合对比表明, 本文提出的植物种群密度变化模型具有较大的的优越性和准确性, 可在一般林分自然稀疏过程研究及生产实践中推广应用。
生境不同的林分所能承受的最大密度有一定差异, 表现在种群密度自我调节强度的不同。生境好, 种群自我调节控制的强度高, 灵敏度强; 生境差时, 调节控制强度低, 灵敏度弱。因此, 要系统全面地反映不同生境条件一般林分自然稀疏过程的密度变化规律, 必须分析本文提出的方程(13)式中有关参数与地位指数的关系, 将地位指数引入方程。例如, 我们用幂函数表达(13)式中的参数a和θ与地位指数SI的关系, 用不同立地的126对固定标准地复测资料拟合(13)式, 求得各参数为:a =3.545SI0.08211, b = 15.5775, c =0.2647, θ=0.192SI0.9256。相关指数R2达0.9725, 说明拟合效果极显著, 可用于模拟不同立地条件下不同生长发育阶段杉木人工林一般林分自然稀疏过程的密度变化规律。
植物种群自然稀疏过程的密度变化规律, 反映的是由各种密度所形成的群体以及组成群体的个体之间相互作用的规律, 但并非任一密度的林分都会发生自然稀疏。例如, 对于人工林, 在造林初期(林分郁闭前)和间伐后(间伐强度足够大)的一段时间内, 林木间拥有较充足的生长空间, 个体树木间的相互作用很小或基本上没有, 林分株数属于等株生长, 不发生自然稀疏。随着林分的进一步生长发育, 林木间开始为争夺生存空间而展开竞争, 且随个体树木的不断增长而竞争加剧, 导致了林分发生自然稀疏而进入自然生长状态。因此, 根据林分平均木(如平均胸径、平均材积)的大小和株数的关系来研究自然稀疏, 应考虑在给定林分平均木大小的情况下, 只有当株数达到足够大时才会发生自然稀疏, 即发生自然稀疏的株数下限, 现有的自然稀疏最大密度法则, 只明确了林分发生自然稀疏过程中所能承受的最大密度, 即上限, 对于下限未予规定, 这是目前研究同龄纯林自然稀疏过程密度变化规律尚未解决的问题, 也是一个难点, 即尚无准确判断林分是属于等株生长还是自然生长的标准, 因此, 今后应加强这方面的研究。
方精云. 1995. 一种描述植物种群自然稀疏过程的经验模型. 林业科学, 31(3): 277-282. |
李凤日, 蒋伊尹, 刘兆刚. 1997. 广义Korf生长方程的推导及其应用. 林业资源管理, (特刊): 212-217. |
刘金福, 洪伟, 李茂瑾. 1998. 格氏栲种群调节模型研究. 热带亚热带植物学报, 6(4): 309-314. |
唐守正. 1993. 同龄纯林自然稀疏规律的研究. 林业科学, 29(3): 234-241. DOI:10.3321/j.issn:1001-7488.1993.03.005 |
张大勇, 赵松林. 1985. 森林自疏过程中密度变化规律的研究. 林业科学, 21(4): 369-374. |
只木良也. 1963. 竞争密度效果な基にした干材积收获予测. 林业科学, 15(4): 1-19. |
Hozumi K. 1977. Ecological and mathematical considerations on self-thinning in ever-aged pure standsⅠ, Mean plant weight-density trajectory during the course of self-thinning. Bot.Mag. Tokyo, 90: 165-179. DOI:10.1007/BF02488355 |
Hozumi K. 1983. Ecological and mathematical considerations on self-thinning in ever-aged pure stands Ⅲ, Effect of the linear growth factor on self-thinning. Bot. Mag. Tokyo, 96: 171-191. DOI:10.1007/BF02498999 |
John P, et al. 1994. Stand and tree dynamics of uneven-aged ponderosa pines. For. Sci., 40(2): 289-302. |
J L克拉特, J C弗尔, C H布雷斯特等著; 范济洲, 董乃钧, 于政中等译.用材林经理学---定量方法.北京: 中国林业出版社, 1987, 101~106, 127~130
|
Yoda K, et al. 1963. Self-thinning in overcrowded pure stands under cultivated and natural conditions. J Biol Osaya City Univ, 14: 107-129. |