 2001, Vol. 37
2001, Vol. 37文章信息
- 杨玉盛, 陈光水, 何宗明, 陈银秀, 谢锦升.
- Yang Yusheng, Chen Guangshui, He Zongming, Chen Yingxiu, Xie Jingsheng.
- 杉木观光木混交林群落细根净生产力及周转
- NET PRODUCTIVITY AND TURNOVER RATE OF FINE ROOTS IN MIXED FOREST OF CUNNINGHAMIA LANCEOLATA AND TSOONGIODENDRON ODORUM
- 林业科学, 2001, 37(专刊1): 35-41.
- Scientia Silvae Sinicae, 2001, 37(专刊1): 35-41.
-
文章历史
- 收稿日期:2001-01-08
-
作者相关文章


2. 福建农林大学林学院 南平 353001
2. Forestry College of Fujian Agriculture and Forestry University Nanping 353001
杉木(Cunninghamia lanceolata)人工纯林多代连栽地力衰退问题已引起人们的极大关注,而杉阔混交林则被认为是维持杉木人工林长期生产力的较好途径之一(俞新妥,1996;盛炜彤,1992;杨玉盛,1998)。福建林学院莘口教学林场从60年代起,就已结合营林生产开展了格氏栲(Castanopsis kawakamii)、木荚红豆树(Ormosia xylocarpa)、观光木(Tsoongiodendron odorum)、建柏(Fokienia hodginsii)等珍贵树种的人工造林试验研究,本文对1973年营造的现已有27 a生的杉木观光木混交林的生产力、养分循环、能量流动及土壤肥力演变等进行了系统研究,为揭示杉观混交林的结构与功能提供帮助。本文为该项目的一部分。
1 试验地概况试验地位于福建三明福建林学院莘口教学林场小湖工区(北纬26°11′30″,东经117°26′00″),属中亚热带季风型气候,年均气温19.1℃,年均降水量1749 mm,年均蒸发量1585.0 mm,年均相对湿度为81%,无霜期300 d左右,土壤是由砂页岩发育的红壤。1973年用实生苗造林,初植密度为3000株·hm-2,混交林为行间混交(杉木与观光木比例为3:1)。杉木纯林现保留密度为1100株·hm-2,平均树高(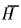

1999-01分别在混交林和纯林中各设立3块20 m×20 m固定标准地,从1999-01至2000-01隔月于月底用内径6.8 cm的土钻在各林分每块标准地的上中下部随机钻取土芯10个,每个林分共30个,深度为1 m。取出土芯并用流动水浸泡、漂洗、过筛,拣出根系,用放大镜、剪刀、镊子等工具分出各目的树种根和林下植被根,捡出直径<2 mm的细根,并根据根系外形、颜色、弹性、根皮与中柱分离的难易程度来区分活死根,对目的树种细根进一步细分成3个径级(1~2 mm、0.5~1 mm、<0.5 mm),将各根样品置于80℃烘箱中烘干至恒重后称重,并按以下公式计算细根生物量:

|
在样地附近从表层土(0~20 cm)中随机收集目的树种(细分成3个径级)和林下植被的活根与死根的混合样品,自然风干后称取5 g样品装入18 cm×18 cm、孔径为0.25 mm的尼龙网袋中,每袋样品均换算为干重,埋入近地表 10 cm处,放置样地中部,每个径级共100袋,以后每隔一定时间从各点中随机抽取各径级样品各6袋,用于失重和化学成分分析。
2.3 细根净生产力和周转应用极差公式计算(McClaugherty et al., 1982),公式如下:

|
式中,M、P、D、T分别代表细根年死亡量、年生长量、年分解量和周转率;Mmax、Mmin分别为死细根现存量最大值和最小值;Pmax、Pmin和Y分别表示活细根现存量的最大值、最小值和平均值。
3 结果与分析 3.1 群落细根数量及其动态 3.1.1 群落细根现存量由表 1可见,中亚热带杉木观光木混交林<2 mm细根的现存量为5.381 t·hm-2,比杉木纯林(4.584 t·hm-2)增加17.4%,高于会同的杉木和火力楠纯林与杉木火力楠混交林及鼎湖山的针阔叶混交林,但低于鼎湖山的季风常绿阔叶林、武夷山的甜槠林和长白山的阔叶红松林(廖利平等,1995;温达志等,1999;李凌浩等,1998)。杉木和观光木混交后群落细根的数量增加,有利于其吸收土壤养分和水分,从而促进混交林群落中林木的生长,这与廖利平等在会同对杉木火力楠混交林和杉木纯林的研究相似(廖利平等,1995)。混交林中杉木的细根现存量与观光木的现存量之比为4.69:1,大大高于这两个树种的株数比(2:1),表明杉木单株的细根量比观光木的大(表 1),这可能亦是混交林中杉木生长量大于观光木的主要原因之一。混交林和纯林林下植被细根(包括灌木与草本)分别占群落细根总量达14.1%和19.0% (表 1),说明林下植被层细根占有较大比重,不宜忽视。从死活根生物量所占比例来看,杉木和观光木各径级活死细根比例均约略为7:3,这与鼎湖山季风常绿阔叶林和针阔叶混交林的活死细根比例相似(70.1:29.9;65.6:34.4) (温达志等,1999),表明不同林分类型或不同径级活死细根比例均相对稳定。杉木、观光木<0.5 mm细根占<2 mm细根量的60.5%~64.2%,说明<0.5 mm细根是乔木细根库的主体。
|
|
在森林生态系统中,与地上部分叶片承担着光合作用的功能相配合,林木细根担负着吸收土壤中水分和养分的功能,对森林生态系统生长发育起着重要的作用;与地上部分每年大量叶片凋落同时长出大量新叶相似,细根在每年枯死同时亦长出大量新根,因此枯死细根与地上部分凋落物对森林生态系统养分循环等均起着重要的作用(Vogt et al., 1983)。杉木活细根现存量变化呈双峰型,在1月份较小,3月时出现第1次峰值,随后下降,7月出现低峰,9月又达到另一峰值,后又呈下降趋势(图 1)。而观光木活细根现存量动态变化与杉木的略有不同,两次峰值出现时间亦为3月和9月,而两次低值出现时间则为5月和1月(图 1)。杉木死细根现存量变化呈单谷型,1月最大,后逐渐下降,5月为一年中的最低值,而后又不断增加,至1月达最大值(图 1)。观光木死细根一年中的最低值出现在3月(图 1)。这与鼎湖山、长白山等地的研究结果不尽相同。鼎湖山季风常绿阔叶林和针阔混交林活细根7月最大,12月最低,而死细根4月或7月最大,12月最小(温达志等,1999);长白山阔叶红松林活细根生物量最低值出现在6月,最大值出现在10月,而死细根最低值在6月,最高值出现在9月(单建平等,1993)。细根现存量动态差异的原因与树种生物学特性和各地的气候差异有关。本试验地处中亚热带,1月气温较低,活细根量较低,且死细根分解较慢,经过一个冬季的积累,死细根量较高;早春时林地土温回升快,降雨量较多,土壤湿润,有利于根系大量生长,3月细根量出现第1次高峰,同时,残留的死细根分解速率加快,死细根量开始减少;随着气温的继续上升和地上部分生长速度快速增大,光合作用产物大部分用于维持地上部分生长之需,分配给细根的数量大量减少,同时,细根供应地上部分生长所需的水分和养分数量急剧增加,亦导致部分细根的死亡,因此活细根数量呈减少趋势,另一方面死细根的分解速率加快,死根量最低;至7月,高温干旱,林木生长量较小,特别是土壤含水量下降,导致细根大量死亡,活细根出现极低值,而死细根量则明显上升;此后由于降雨量增加,林木生长量加大,活细根量有所增加,至9月活细根量出现另一高峰,而死根量也仍在上升;随后,由于气温开始降低,林木生长减慢,活细根量不断减小,由于细根死亡增加和死细根分解速率减慢,引起死细根量的不断累积,至1月时,活细根量出现低值,而死根量达最大值。混交林杉木与观光木细根变化动态差异的原因主要可能是杉木和观光木对水分和养分利用方式差异有关。林下植被细根现存量的变化与杉木的相似(图 2)

|
图 1 乔木层细根生物量现存量动态变化 Fig. 1 Dynamics changes in existing amount of fine roots biomass in the tree stratum 混交林杉木C.lanceolata in mixed forest: ◆活细根Living roots, ◇死细根Dead roots;观光木T.odorum: ▲活细根Living roots, △死细根Dead roots;纯林杉木C.lanceolata in pure forest: ●活细根Living roots, ○死细根Dead roots. |

|
图 2 林下植被细根生物量现存量动态变化 Fig. 2 Dynamics changes in biomass of existing fine roots in the undergrowth 混交林Mixed forest: ◆活细根Living roots, ◇死细根Dead roots;纯林Pure forest: ▲活细根Living roots, △死细根Dead roots. |
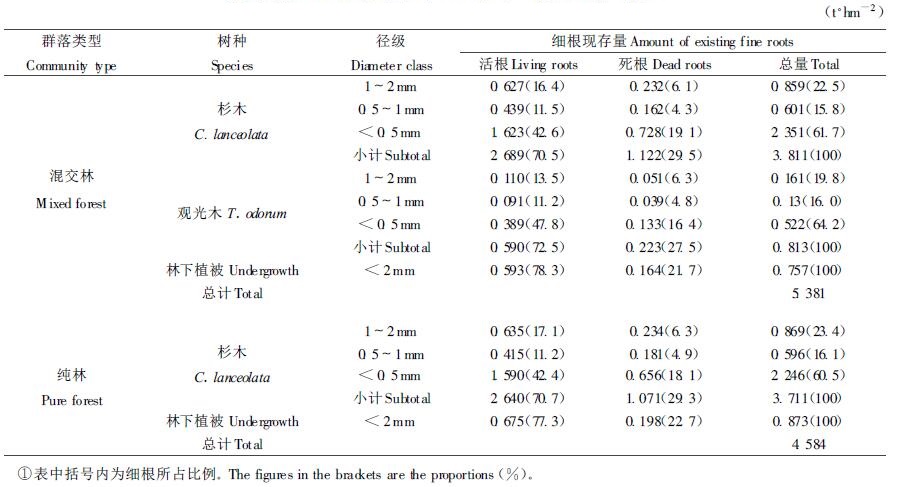
细根的周转在森林生态系统中起着重要的作用(李凌浩等,1998)。观光木细根分解第1年后的失重率高于杉木的,而同一树种不同径级细根的失重率又随径级的减小而增大(表 2)。本文杉木和观光木细根分解的失重率分别与会同的杉木和火力楠的相近(廖利平等,1995),且比各自树种的凋落物分解率要快得多(未刊资料)。由表 2也可见,林下植被细根的失重率比乔木层细根的高,说明林下植被细根的分解速率快于乔木层细根,且杉木纯林林下植被细根的失重率略高于混交林林下植被细根,这与纯林林下植被细根中草本细根所占比重较高有关。
|
|
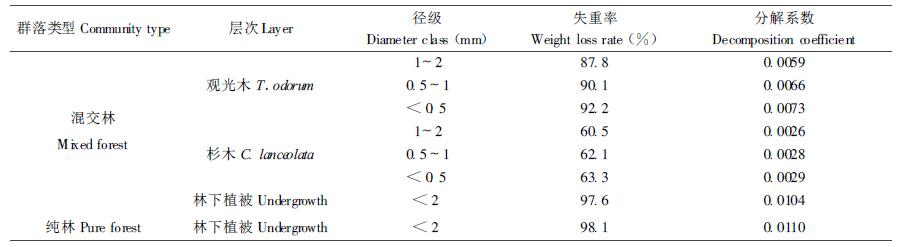
根据一年中每隔2个月共6次取样数据,采用下式:X/X0=exp (-kt) (其中,X0为分解初始干重,X为分解t d后的残留物干重,k为分解系数) (Olson, 1963),建立细根不同径级指数方程(相关系数均达0.80以上),求得分解系数k (表 2)。由表 2可见,林下植被细根的分解系数k值大于乔木层的,而观光木的k值又大于杉木的,且同一树种k值随径级的减小而增大。
3.3 细根的年生长、死亡和周转率利用细根的第1年失重率乘以死细根年均生物量现存量得细根年分解量,再利用McClaughery (1982)提出的极差模型计算年生长量、年死亡量和周转率得表 3。
|
|

从表 3可见,杉木和观光木不同径级细根的年分解量、年死亡量和年净生产力均为:<0.5 mm>1~2 mm>0.5~1 mm,表明<0.5 mm的细根是群落中细根周转的主体。混交林和纯林细根年死亡量分别为2.119和1.894 t·hm-2,分别是相应群落地上部分凋落物量(6.759和6.804 t·hm-2)的31.4%和27.8%,其中混交林杉木、观光木和纯林杉木细根的年死亡量分别为1.461、0.348和1.519 t·hm-2,而它们的年凋落物量分别为4.182、1.086和5.021 t·hm-2 (未刊资料),细根年死亡量分别占其年凋落物量的34.9%、32.0%和30.3%,这比长白山阔叶红松林细根死亡量占凋落物比重约60%低(单建平等,1993),但比鼎湖山常绿阔叶林细根(<5 mm)年死亡量占其年凋落物量的24.1%高(温达志等,1999),这可能与林木的年龄和生物学特性有关。但由于细根的分解速率比凋落物快,且细根的养分含量比凋落物的高,因而细根在群落养分年归还中的作用可能比凋落物的大(廖利平等,1995)。混交林和纯林林下植被细根的年死亡量亦分别占群落细根死亡量的10.9%和15.6%。
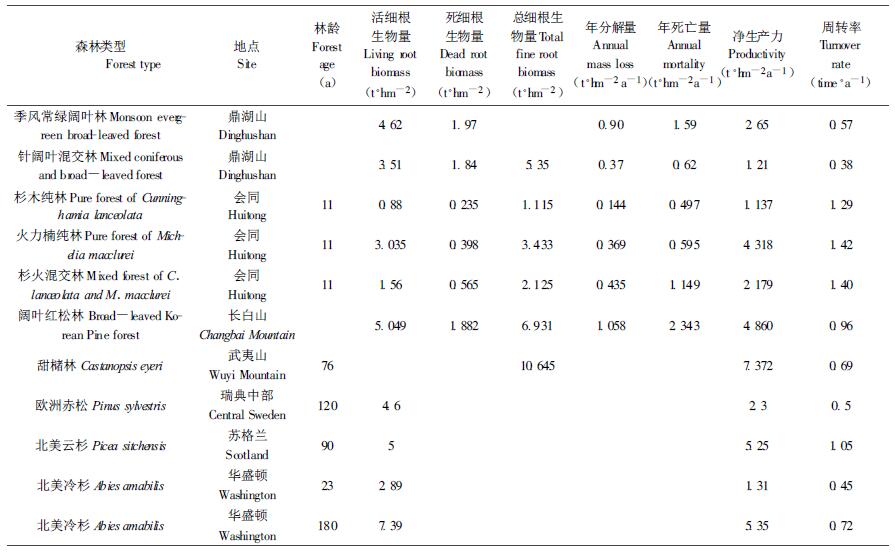
混交林细根的年净生产力达4.124 t·hm-2a-1,是纯林的1.17倍(表 3);与同类研究结果相比,本文中细根年净生产力均处于已报道的研究结果范围内(1.137~7.372 t·hm-2a-1) (表 4)。混交林和纯林的细根生产力分别占各自群落净第一性生产力(18.003和16.846 t·hm-2a-1,未刊资料)的22.9%和20.9%,高于长白山阔叶红松林细根所占的比例(19.4%) (单建平等,1993),亦高于鼎湖山的季风常绿阔林和针阔中混交林细根(<5 mm)所占的比例(16.8%和17.4%) (温达志等,1999),更远高于细根占各自群落总生物量(254.906和200.605 t·hm-2)的比例(2.1%和2.3%)。
|
|
细根的周转与细根的吸收需要有关,即树木必须经常在新的部位生长新根代替旧根以适应土壤中不断变化的水分和养分状况,以达到充分吸收水分和养分的同时避免过多的无用根系的呼吸消耗(Bartsch, 1987)。单建平等则认为树木的地下部分也像其他地上部分一样有老化、死亡、更替的过程(单建平等,1992)。混交林中观光木各径级细根的年平均周转速率为1.16,比杉木(0.96)快,同一树种的不同径级细根的年周转速率随径级的减小而增大,这与随径级减小细根的木质化程度低、分解快有关。林下植被细根的周转率均大于各自的乔木层细根,这与林下植被细根的木质化程度比乔木层细根低有关。混交林中杉木细根的周转速率稍快于纯林杉木的,这与混交林杉木<0.5 mm的细根所占比例稍高于纯林杉木的有关。混交林群落细根的年周转率(1.07)亦比纯林的(1.06)稍高(表 3),说明杉观混交27 a后,不但其细根净生产力大,而且加速了细根的周转。
从表 3还可见杉木观光木的细根周转率在0.89~1.18之间,落在多数已报道0.5~1.20次·a-1的范围内,但处于其上限,说明中亚热带的杉木混交林及杉木纯林细根周转速率是较大的(表 4)。
本研究的细根周转率高于鼎湖山的季风常绿阔叶林和针阔叶混交林(温达志等,1999)、武夷山的甜槠林(李凌浩等,1998)、瑞典的欧洲赤松(Person, 1983)和华盛顿的北美冷杉(Vogt et al., 1983),而与长白山阔叶红松林(单建平等,1993)和苏格兰北美云杉(Deans, 1981)相当,但低于会同的杉木纯林及混交林(廖利平等,1995) (表 4)。造成细根周转率差异的原因,与研究地所处地区的气候条件、树种特性、林龄、干扰状况等有关。由于本研究地处中亚热带,气候温暖湿润,水热条件较好,树种皆为速生树种,细根的生长与更替较频繁,因而细根周转速率普遍比温带树种及天然林(树龄较大)大。而会同杉木火力楠混交林与本研究地所处气候条件相似,但其年龄(10 a生)比本研究的(27 a生)小,林木正处于快速生长期,因而细根周转速率比本研究的大(廖利平等,1995)。
4 小结(1) 27 a生杉木观光木混交林和杉木纯林群落的细根生物量仅分别占群落总生物量的2.1%和2.3%,而其细根的年净生产力却占各自群落第一性生产力的22.9%和20.9%,表明细根在群落的净生产力组成中占有重要地位。
(2) 混交林和纯林细根的年死亡量分别为2.119 t·hm-2和1.894 t·hm-2,是各自群落地上部分凋落物量的31.4%和27.8%,表明细根枯死亦是群落向土壤子生态系统归还有机质的重要途径之一。
(3) 乔木层细根的周转速率大小顺序为混交林中观光木>混交林中杉木>纯林杉木,说明阔叶树细根周转速率大于针叶树的;而林下植被层细根周转速率均高于相应的乔木层,表明林下植被层细根在群落细根周转中占有重要地位。
(4) 杉观混交林细根的生物量、年死亡量、年生产力、年周转率均高于纯林,表明杉木与观光木混交后,促进了群落细根的生长,提高群落细根的周转率,这对林地土壤具有较好的改良作用,从而促进群落中杉木的生长。
单建平, 陶大立, 王森, 等. 1993. 长白山阔叶红松林细根周转的研究. 应用生态学报, 4(4): 241-245. |
单建平, 陶大立. 1992. 国外对树木细根的研究动态. 生态学杂志, 11(4): 46-49. |
李凌浩, 林鹏, 邢雪荣. 1998. 武夷山甜槠林细根生物量和生长量研究. 应用生态学报, 9(4): 337-340. DOI:10.3321/j.issn:1001-9332.1998.04.001 |
廖利平, 陈楚莹, 张家武, 等. 1995. 杉木、火力楠纯林及混交林细根周转的研究. 应用生态学报, 6(1): 7-10. DOI:10.3321/j.issn:1001-9332.1995.01.005 |
盛炜彤主编. 1992. 人工林地力衰退研究. 北京: 中国科学技术出版社.
|
温达志, 魏平, 孔国辉, 等. 1999. 鼎湖山南亚热带森林细根生产力与周转. 植物生态学报, 23(4): 361-369. DOI:10.3321/j.issn:1005-264X.1999.04.009 |
杨玉盛. 1998. 杉木林可持续经营的研究. 北京: 中国林业出版社.
|
俞新妥主编. 1996. 杉木栽培学. 福州: 福建科学技术出版社.
|
Bartsch N. 1987. Responses of root systems of young Pinus sylvestris and Picea-abies plants to water deficits and soil acidity. Can. J. For. Res., 17: 805-815. DOI:10.1139/x87-128 |
Deans J D. 1981. Dynamics of coarse root production in a young plantation of Picea sitchersis. Forestry, 54: 139-155. DOI:10.1093/forestry/54.2.139 |
McClaugherty C A, Aber J D. 1982. The role of fine roots in the organic matter and nitrogen budgets of two forested ecosystems. Ecology, 63: 1481-1490. DOI:10.2307/1938874 |
Olson J S. 1963. Energy storage and the balance of producers and decomposers in ecological systems. Ecology, 44: 323-331. |
Person H. 1983. The distribution and productivity of fine roots in boreal forests. Plant and Soil, 71: 87-101. DOI:10.1007/BF02182644 |
Vogt K A, Vogt D J, Asbjornsen H. 1983. Production, turnover and nutrient dynamics of above-and below-ground detritus of world forests. Advances in Ecological Research, 15: 303-378. |