 2001, Vol. 37
2001, Vol. 37文章信息
- 罗世家, 邹惠渝, 梁师文.
- Luo Shijia, Zou Huiyu, Liang Shiwen.
- 黄山松与马尾松基因渐渗的研究
- STUDY ON THE INTROGRESSIVE HYBRIDIZATION BETWEEN PINUS HWANGSHANENSIS AND P. MASSONIANA
- 林业科学, 2001, 37(6): 118-122.
- Scientia Silvae Sinicae, 2001, 37(6): 118-122.
-
文章历史
- 收稿日期:2000-05-22
-
作者相关文章


2. 南京林业大学 南京 210037;
3. 安徽黄山风景区园林局 黄山 242709
2. Nanjing Forestry University Nanjing 210037;
3. Landscape Bureau of Mt.Huangshan, Anhui Province Huangshan 242709
渐渗是指一个种的遗传物质通过杂交与反复回交穿越种间障碍转入到另一个物种内的现象, 又称渐渗杂交(Introgressive Hybridization) (Anderson, 1953)。松属是一个种间杂交比较容易的属。关于松属的种间杂交国内外已有不少报道。而关于黄山松与马尾松的自然杂交, 却一直存有争议。黄山松分布于我国东部中亚热带高山, 海拔700 m以上。马尾松广布于我国亚热带地区。在长江下游地区, 垂直分布于海拔700 m以下, 长江中上游地区, 其垂直高度可达1200 ~ 1500 m左右。在黄山松与马尾松共同分布地区, 海拔700 ~ 1000 m之间形成了一个黄山松与马尾松垂直分布的重叠区。这一区间内的许多松树生长高大、通直, 其形态特征或多或少介于二者之间(邢有华等, 1992), 以安徽黄山的云谷寺、慈光阁一带最富有代表性。对这类松树的处理, 目前存有异议。李书春在江西武宁观察到类似的现象, 认为这类松树仍与黄山松关系比较近, 而发表为黄山松的一个新变种——杉松(Pinus hwangshanensis Hsia var. wulingensis S. C. Li) (李书春, 1981)。祁承经、林亲众根据湖南的标本, 认为其形态特征与黄山松极其相似, 但树脂道为边生绝非中生而与马尾松关系较近, 将其作为马尾松的一个新变种——武陵松(P. massoniana Lamb. var. wulingensis C. J. Qi et Q. Z. Lin) (祁承经等, 1988)。陈忠毅根据采自黄山的材料, 利用Anderson杂种指数法分析表明黄山松与马尾松之间存在天然杂交和基因渐渗现象。将形态特征介于马尾松与黄山松之间的类群称之为渐渗群体, 渐渗群体的树脂道全为兼生, 而认为是天然杂种, 但渐渗群体的平均指数和它的总值偏向于黄山松, 因而认为是天然杂种与黄山松反复回交的结果(陈忠毅, 1986)。
本文采用随机扩增多态性DNA(RAPD)遗传标记法研究黄山松与马尾松的渐渗杂交。
1 实验材料RAPD标记法为显性标记, 不能区分基因的纯合性与杂合性, 而裸子植物的胚乳为单倍体, 因此本实验取黄山松与马尾松的单倍体胚乳作为实验材料。
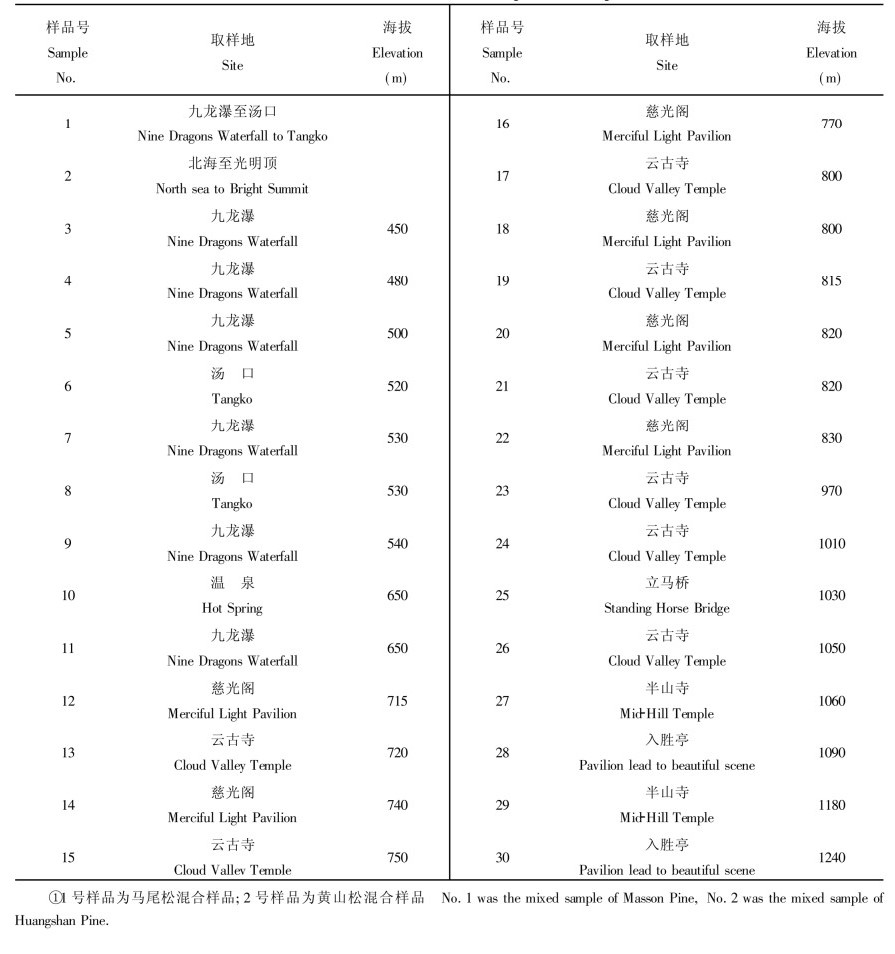
材料采自安徽黄山风景区, 根据海拔高度和形态特征将材料分为:马尾松、黄山松, 以及形态特征介于马尾松与黄山松之间, 海拔高度600 m以下、600 ~ 800 m、800 ~ 1000 m、1000 ~ 1200 m等6个组。马尾松采自九龙瀑至汤口一带, 黄山松采自北海至光明顶一带, 海拔均在1200 m以上, 其余4组的材料沿九龙瀑—云古寺—白鹅岭和慈光阁—半山寺—玉屏楼两条线路采集。每个组取10株不同个体的饱满种子各1粒, 剥去种皮和胚, 取其胚乳用SDS提取液提取DNA(李宽钰, 1996)。
将马尾松组和黄山松组各个个体的DNA按照等浓度分别混合。其余4组各取7个样品, 共计30个DNA样品(表 1)
|
|
DNA扩增反应用美国生产的PEKIN9600 PCR仪, 用Operon Tech.公司生产的引物480个, 以任意一个DNA样品为模板对引物进行初次筛选, 获得扩增效果较好的引物30个, 然后以马尾松和黄山松为模板对30个引物进行多态性筛选, 共获得在马尾松与黄山松之间具有稳定多态的引物10个, 用这10个引物, 分别对30个DNA样品进行RAPD扩增。
3 结果分析10个引物, 共检测到29个多态位点, 其中11个位点为马尾松有扩增产物而黄山松没有扩增产物的位点, 9个位点为黄山松有扩增产物而马尾松没有扩增产物的位点。5个位点为黄山松与马尾松均没有扩增产物, 为渐渗群体中出现的多态位点。4个位点为黄山松与马尾松都有扩增产物而渐渗群体中有分离的位点。29个位点在6个组中的频率见表 2。
|
|

从表中可以看出, 位点3、15、19等3个位点随着海拔高度的升高, 其频率逐渐增加。位点4、6、9、10、16、17、22等7个位点随海拔的升高其频率逐渐降低, 说明黄山松与马尾松之间存在基因渐渗现象, 而且马尾松的基因向杂交群体中渐渗的频率高于黄山松。部分位点频率与海拔的关系见图 1。

|
图 1 基因位点的频率与海拔的关系 Fig. 1 The relation between the frequency of loci and elevation
—◆ —位点3 Locus 3;—■—位点15 Locus 15; 位点6 Locus 6; —●—位点10 Locus 10; 位点6 Locus 6; —●—位点10 Locus 10; 位点19 Locus 19; 位点19 Locus 19;  位点4 Locus 4. 位点4 Locus 4.
|
将渐渗群体各个体(3 ~ 30号样品)的扩增谱带与马尾松和黄山松对比, 判定各个体同马尾松与黄山松的相同位点数, 统计结果见表 3。
|
|

从各个个体的扩增结果来看, 9、16、21号个体与马尾松相同的位点较多, 没有或很少有与黄山松相同的位点, 属于马尾松种群。26、27、28、29、30号个体均采自海拔1050 m以上, 它们与黄山松相同的位点明显多于与马尾松相同的位点, 属于黄山松种群。12个个体扩增出13个位点, 其中7个位点与马尾松相同, 6个与黄山松相同, 18号个体扩增出12个位点, 其中6个位点与黄山松相同, 另6个位点与马尾松相同, 23号个体扩增出10条带, 其中与黄山松与马尾松相同的位点数均为5, 可推断12、18、23等个体为黄山松与马尾松的天然杂种, 与位点比例与之比较相似的22号、24号等个体一起, 代表了杂交种群。另外随着海拔高度的不同, 以及各取样材料所处的世代不同, 与马尾松或黄山松回交频率的差异, 形成了一些梯度类型。
将RAPD扩增结果, 运用Popgen16软件计算出各样品之间的遗传距离, 用不加权平均距离法(UPGM)进行聚类分析, 结果见图 2。

|
图 2 聚类分析树状图 Fig. 2 The dendrogram of clust analysis |
所有材料分成了5个类群。9、10、16、21号个体的位点主要与马尾松相同。它们在遗传距离的聚类图上, 恰好与1号马尾松聚成一类(类群Ⅰ为马尾松种群)。与黄山松具有较多相同位点的26、27、28、29、30号个体均采自海拔1050 m以上, 在遗传距离聚类图上也正好与2号黄山松聚成一类(类群Ⅱ为黄山松种群)。12、18、23号个体与黄山松和马尾松相同的位点比为1: 1, 与位点比例与之比较接近的24号、17号、22号个体一起组成了聚类图上的类群Ⅲ, 代表了杂交类群。类群Ⅳ和类群Ⅴ为杂交种与一方或双方反复回交, 形成的一些渐渗类群。
4 结论各位点在不同海拔高度的频率, 各实验样品所具有的与马尾松和黄山松相同的位点数以及遗传距离的聚类分析均表明黄山松与马尾松在垂直分布重叠区域内存在自然杂交与基因渐渗现象, 而且马尾松的基因向杂交群体中渐渗的频率高于黄山松。
黄山松与马尾松的自然杂交和基因渐渗, 可能与我国东南部第四纪地质与地貌的变化有关。由于冰期气温的逐步下降, 使黄山松降至华中平原(徐炳声, 1956); 间冰期, 由于气温的转暖, 黄山松向中亚热带高山侵移, 形成目前在我国东部中亚热带高山的岛屿状分布。同时, 由于间冰期气温的回升, 马尾松向高位山地侵移, 原本有地理隔离的两个种之间的地理隔离被打破, 使两个种之间的杂交与渐渗得以发生。
陈忠毅.安徽黄山松与马尾松的天然杂交.中国科学院华南植物研究所集刊, 1986, 第二集: 38~ 40
|
祁承经, 林亲众. 1988. 马尾松一新变种. 植物研究, 3(3): 143-145. |
邢有华, 方永鑫, 吴根荣. 1992. 安徽大别山黄山松与马尾松天然杂交的初步研究. 安徽林业科技, (4): 5-9. |
李书春. 1981. 黄山松一新变种——杉松. 安徽农学院学报, (1): 39-40. |
李宽钰. 1996. 白杨派、青杨派和黑杨派的DNA多态性及系统进化的研究. 南京林业大学学报, 20(1): 6-11. |
李宽钰. 1997. 用RAPD探讨毛白杨起源. 植物学学报, 35(1): 24-31. |
徐克学.科学的分类方法——数量分类学.生物科学参考资料, 北京: 科学出版社, 1983, (第十七集): 16~ 25
|
徐克学, 李德中. 1983. 我国人参属的数量分类初试. 植物分类学报, 21(1): 34-41. |
钟扬, 等. 1990. 数量分类的方法与程序. 武汉: 武汉大学出版社.
|
邹喻苹. 1995. RAPD分子标记简介. 生物多样性, 3(2): 104-108. DOI:10.3321/j.issn:1005-0094.1995.02.010 |
Anderson E. 1953. Introgresive hybridization. Biol.Rev, 28: 280-307. |
Garcia G M, Stalker H T, Kochert G. 1995. Introgression analysis of an interspecific hybrid population in peanuts (Arachis hypogaea L.) Using RFLP and RAPD Markers. Genome, 38: 166-176. DOI:10.1139/g95-021 |
Keim P, et al. 1989. Genetic analysis of an interspeciefic hybrid swarm of Populus occurrence of unidirectional intogression. Genetics, 123: 557-565. |
Smous P E, Salor L C. 1973. Studies of the Pinus rigida-serotina complex Ⅱ. Natural hybridization among the Pinus rigida-serotina complex, P. taeda and P. echinata. Ann. Missouri Botanical Gardens, 60(2): 192-203. DOI:10.2307/2395085 |