 2001, Vol. 37
2001, Vol. 37文章信息
- 张康健, 白明生, 张檀, 马希汉, 高锦明.
- Zhang Kangjian, Bai Mingsheng, Zhang Tan, Ma Xihan, Gao Jinming.
- 杜仲叶次生代谢物与个体生长发育特性的研究
- A STUDY ON THE GROWTH FEATURES OF INDIVIDUALS AND SECONDARY METABOLITES OF EUCOMMIA ULMOIDES LEAVES
- 林业科学, 2001, 37(6): 45-51.
- Scientia Silvae Sinicae, 2001, 37(6): 45-51.
-
文章历史
- 收稿日期:1999-07-23
-
作者相关文章


杜仲(Eucommia ulmoides Oliv)是我国特有的经济林树种, 既是贵重的药材, 也是提取天然橡胶的工业原料。杜仲的化学成分很复杂, 近几年来, 国内外研究认为绿原酸、京尼平甙酸、京尼平甙、桃叶珊瑚甙、总黄酮等次生代谢物是杜仲叶中包含量较高的生物活性物质(尉芹等, 1995; 高锦明等, 1999), 即药用有效成分; 杜仲胶是工业开发利用的天然橡胶, 目前已开发出系列产品(张康健等, 1997)。所以, 为了充分开发利用杜仲资源, 提高其经济效益, 选择富含上述次生代谢物的优良个体成为亟待解决的问题。
杜仲在长期的演化过程中, 由于天然杂交、自然选择、人工培育, 并在所分布区内, 长期适应于其所生存的环境条件, 所以往往在形态、生理和遗传特性上产生分化, 表现出个体生长发育的差异, 而这种差异正是导致中草药品质优劣的原因所在。清代的《药性变迁论》中就曾提到“以古方所用之药……今依方施用, 竟有应有不应, 其何故哉?盖有数端焉:一则地气之殊也! ……一则天生与人力之异也。”(林世俊, 1985), 从而说明生态环境引起药用植物种内变异, 并进而影响其药效。许多学者虽然曾对杜仲进行了不少研究(张康健等, 1996; 1997), 但有关这方面的研究却未见报道。故本文对杜仲个体生长发育特性与其叶中次生代谢物(有效成分)含量进行相关分析, 企图为杜仲的良种选育提供表型参考依据。
1 研究材料与方法本实验所用杜仲叶采自原西北林学院杜仲优树汇集圃, 为同一立地条件下生长的、同一年龄(8龄)的40个杜仲无性系(从四川、湖南、贵州、陕西等省选择的优良单株, 通过无性繁殖建立的优树汇集圃)。每一个无性系中, 随机抽取2株, 于1997-06-15由树体同一部位(树冠中部)东南西北四个方向分别采集等量叶片, 杀青(采下叶片后, 立即在烘箱内恒温95 ℃烘5 min), 叶片自然风干后备用。
1.1 次生代谢物含量分析方法 1.1.1 总黄酮的提取及测定总黄酮的测定方法采用的是硝酸铝-亚硝酸钠比色法(冯煦等, 1990)。
1.1.2 杜仲胶的提取测定杜肿胶的提取采用改进的碱浸法(马柏林等, 1994)。
1.1.3 绿原酸、京尼平甙、京尼平甙酸、桃叶珊瑚甙的提取测定本试验采用反相高效液相色谱(HPLC)法同时测定了绿原酸、京尼平甙、京尼平甙酸及桃叶珊瑚甙(Takahashi et al., 1988, 张康健等, 1999)。色谱条件:色谱柱: Nova-pak C18(4 μm, 3.9 mm×75 mm); 流动相:甲醇—水—冰乙酸(10: 90: 1);流速: 1 mL·min-1; 检测波长: 232 nm
采用的主要仪器和试剂为754型紫外-可见分光光度计(上海第三分析仪器厂制造); Waters高效液相色谱系统, 包括486紫外检测器, 510型泵(美国Milford); XWT-204型台式自动平衡记录仪(上海大华仪表厂); 桃叶珊瑚甙(和光纯药工业株式会社, 日本); 京尼平甙(和光纯药工业株式会社, 日本); 京尼平甙酸(和光纯药工业株式会社, 日本); 芦丁(上海试剂工厂); 绿原酸(光谱纯) (德国进口);
1.1.4 硝酸还原酶活力测定方法详见张檀等(1994)的论文。根据硝酸还原酶活力, 进行系统聚类, 把40个无性系单株分为硝酸还原酶活力高(≥2.55)和低(< 2.55)两种类型。
1.2 杜仲个体生长发育特性调查 1.2.1 叶面积采用纸样称重法(章文才, 1991)。测算出单叶面积后进行系统聚类, 结果40个杜仲无性系聚类为A、B两类, A类叶面积≥47.50 cm2, B类叶面积 < 47.50 cm2。
1.2.2 芽开绽期观察标准:全树50 %以上芽鳞片开始脱落、露出小叶时为“芽开绽期”。
1998年春季, 每天早晨8~9时观察发芽情况, 从3月22日第1株芽开绽至4月7日全部芽开绽, 历时17 d, 经系统聚类, 3月30日以前芽开绽聚为一类(芽开绽期早), 3月30日以后芽开绽聚为一类(芽开绽期晚)。
1.2.3 性别、叶形、树皮类型调查杜仲为雌、雄异株, 通过母树原始调查记录和开花时观察花的类型, 记录各无性系的性别。叶片成熟时, 目测观察叶片形状, 分为卵形、椭圆形2种类型。树皮类型根据树皮粗糙度(皮孔数、皮孔突出程度, 纵裂纹深浅)分为光皮、粗皮两类(张康健等, 1997)。
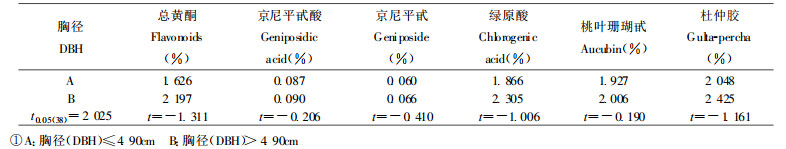
1.2.4 胸径、树高调查胸径调查值经系统聚类, 聚为A、B两类。A类胸径(DBH)≤4.90 cm, B类胸径(DBH)>4.90 cm。树高调查值经系统聚类, 聚为A、B两类。A类树高(Height)≤7.00 m, B类树高(Height)>7.00 m。
2 结果与分析 2.1 不同来源地区和不同无性系次生代谢物含量分析由表 1可见, 其6种次生代谢物含量无论在来源地区间或是无性系间均存在显著差异。
|
|
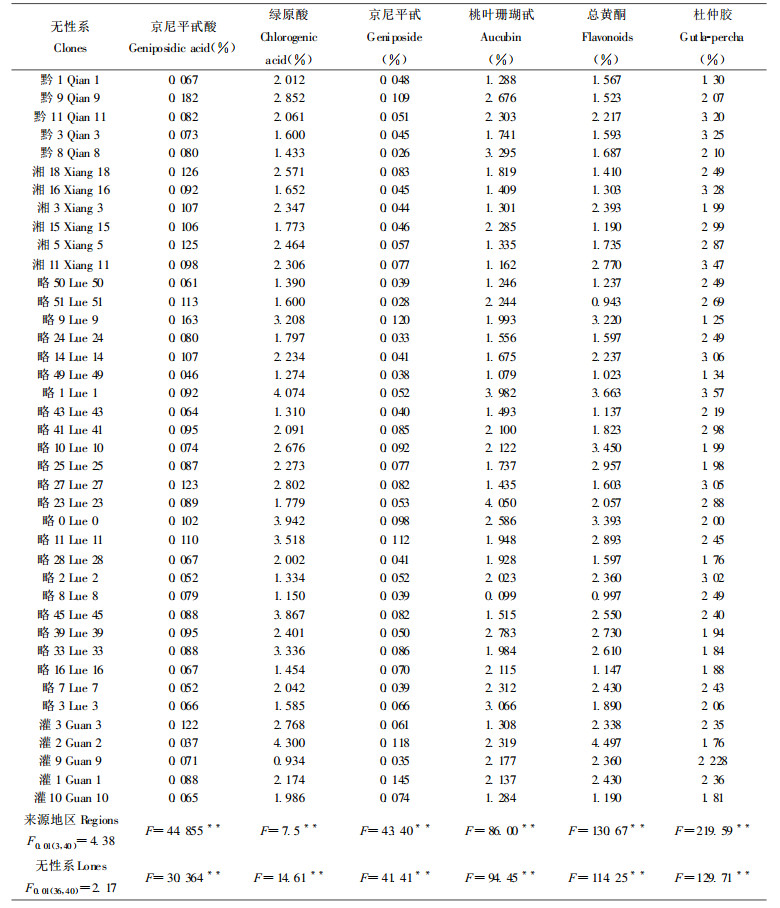
本项研究是在1997年对杜仲叶次生代谢物生长积累动态研究的基础上(张康健等, 1999)进行的, 本文对40个无性系进行了次生代谢物含量的分析, 这40个无性系是从全国杜仲主产区(陕西、贵州、四川、湖南等省)选择的优良单株, 不同的个体其原本所处的生态环境不同, 现在他们虽生活在同一个环境中(原西北林学院杜仲优树汇集圃), 但由于来自不同的生态地理环境, 在原地区多年的天然杂交、自然选择和人工培育的过程中, 已形成了多个在形态结构、生理生化上具有稳定差异的生态类型, 从而引起其次生代谢物含量的差异(见表 1)。因此, 种内的变化不一定是大的外部形态的变化, 在多数情况下表现为表型上的小差异和生理代谢上的差异。
2.2 次生代谢物含量与个体生长发育特性的关系杜仲在漫长的进化过程中, 由于形成了具有一定差异的个体生长发育特性(性状), 以及生理代谢上的差异性, 从而引起次生代谢物含量的差异性。本文就其主要性状(表 2)进行如下分析。
|
|
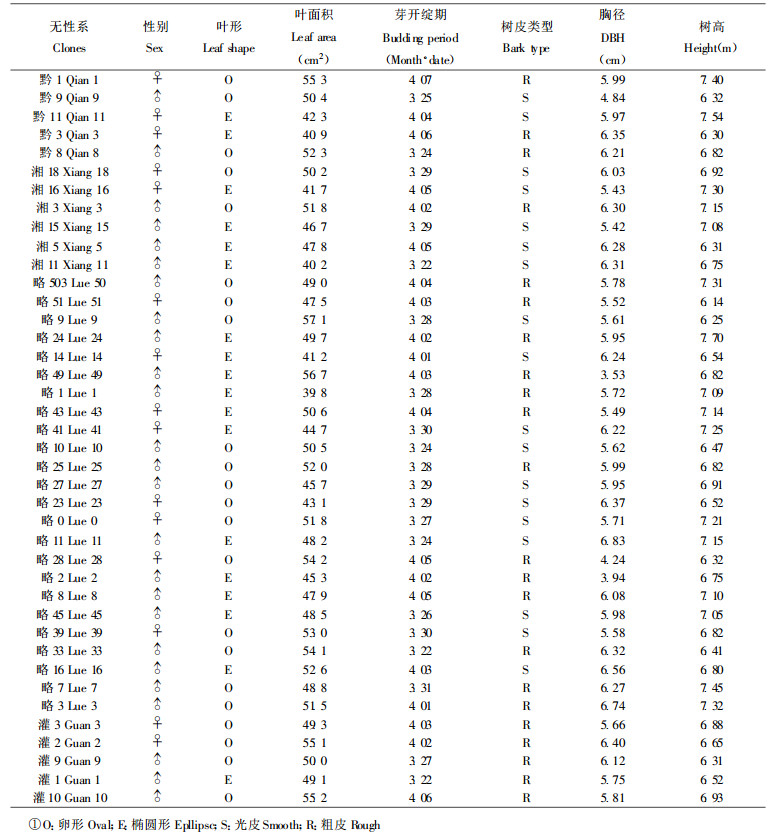
1998年春季, 观测了40个杜仲不同无性系的芽开绽期, 结果表明, 杜仲不同无性系个体间芽开绽期很不相同, 从3月22日第一株芽开绽到4月7日全部芽开绽, 历时17 d。由表 3可见, 芽开绽期早(A)的个体, 其总黄酮、京尼平甙、京尼平甙酸、绿原酸、桃叶珊瑚甙含量显著高于芽开绽期晚(B)的个体, 而杜仲胶的含量无显著差异。
|
|
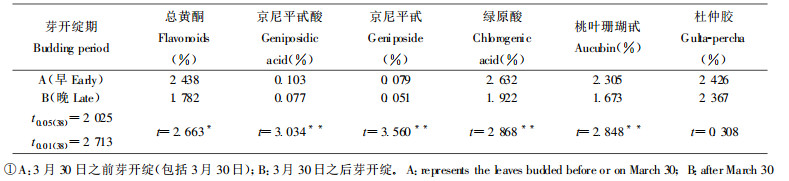
由表 4可看出, 树皮类型对次生代谢物的含量有影响, 光皮类型高于粗皮类型, 但仅对京尼平甙酸、京尼平甙的影响显著, 对其它4种次生代谢物的影响不明显。
|
|
由表 5分析可见, 叶的类型不同, 其次生代谢物含量有差异, 其中卵形叶杜仲胶含量显著地低于椭圆形叶。
|
|
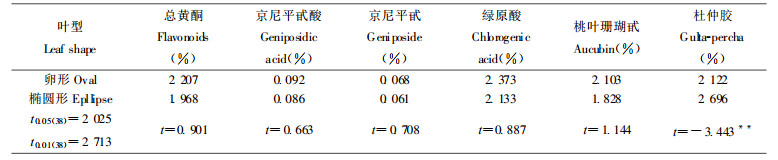
据研究报道(马柏林等, 1996), 杜仲叶中的胶丝与叶脉的网络结构完全一致, 胶丝是叶脉的组成部分。由于卵形叶和椭圆形叶的叶脉有差异, 从而导致其胶丝网络结构出现差异, 影响其杜仲胶含量。
2.2.4 叶面积与次生代谢物含量的相关分析由表 6可见, 杜仲叶中总黄酮、京尼平甙酸、京尼平甙、绿原酸、桃叶珊瑚甙含量在A、B间无显著差异, 即叶面积大小对这5种次生代谢物含量无显著影响; 而杜仲胶的含量却存在极显著差异。故本文着重对杜仲胶含量与其叶面积之间的关系进行回归分析。
|
|

以杜仲胶含量作为因变量(y), 以叶面积作为自变量(x), 建立一元线性回归方程如下: 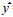

|
图 1 杜仲胶含量与叶面积相关分析散点图 Fig. 1 Correlative plots of Gutta-perch contents and leaf area |
田兰馨等(1983)在研究杜仲叶形态结构时发现, 含胶细胞主要分布在各级叶脉及叶柄维管束的韧皮部。由于杜仲叶大小不同, 其叶脉网络结构的疏密程度不同, 从而导致杜仲胶含量呈现这种规律性的变化。
2.2.5 胸径生长量和树高生长量与次生代谢物含量的关系由表 7、8可见, 胸径生长量、树高生长量大小对次生代谢物含量无显著影响。
|
|
|
|
硝酸还原酶(NR)是植物硝酸盐同化过程中的起始酶, 在氮素代谢中处于关键地位, 据张檀等(1994)报道, NR活力高低反映着植物同化硝酸盐的能力, 一定程度上反映着树木的生长潜力。由此我们对NR活力与次生代谢物含量进行比较分析。
由表 9可见, 不同无性系硝酸还原酶活力的高低对次生代谢物含量影响不明显。
|
|
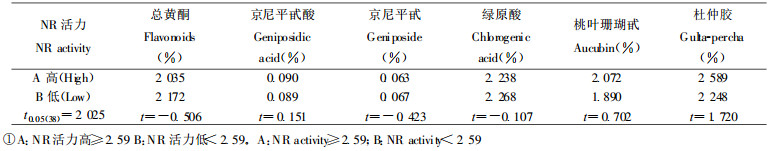
由表 10可见, 雌、雄株差异对杜仲叶次生代谢物含量无影响。
|
|

生物学和生态学研究表明, 40个杜仲无性系的6种次生代谢物含量存在明显的差异; 种内变异是由遗传决定的变异和生态环境引起的表型变异的综合产物(肖小河, 1989)。药用植物都生长在一定的生态地理环境, 在自然选择和人工培育过程中, 其品质变异具有客观必然性。种内变异不一定都产生大的外部形态的变化, 而是在多数情况下表现为生理生化上的变化。遗传因素的多态性和环境条件的多样性都能对植物次生代谢作用产生影响, 而环境因素、遗传因素和表型特征三者之间是相互联系、相互影响的, 致使杜仲不同无性系由于个体生长发育特性的差异而表现出次生代谢物含量的差异。因此, 在研究药用植物的过程中, 要想了解影响品质优劣的根本原因, 就不仅要对其生理特性、化学成分进行研究, 还必须从物种生物学角度对他们的形态学特征、遗传学特征以及生态学特性进行全面系统的综合研究。
杜仲胶含量与叶形明显相关, 且与叶面积呈极显著的负相关(马柏林等, 1996):对杜仲胶丝网络结构进行了研究, 结果表明, 胶丝网络体系与叶片的外形相同, 胶丝网络体系是完全由胶丝组成的一张薄纱, 叶片是由表皮、叶脉和叶肉等组成的器官, 而胶丝的网络结构正好反映了叶脉的网络结构, 这与田兰馨等(1983)关于胶丝分布于叶脉维管束的韧皮部中的结论一致, 本研究结果表明叶形、叶面积与杜仲胶含量关系密切, 其中椭圆形叶杜仲的胶含量明显高于卵形叶, 究其原因, 主要是两种叶形叶脉结构的差异导致其含胶量的差异。再者, 叶开展时, 主要靠居间生长, 增加各部分细胞数目, 叶发育成熟后, 其叶脉网络结构基本相同, 但叶面积不同, 叶脉网络的疏密程度也就不同, 当叶面积较小时, 网络结构相对较密, 所占比例增大, 叶面积较大时则反之, 从而出现了杜仲的胶含量与叶面积呈极显著的负相关现象。因此在选择杜仲高含胶量的优良无性系时, 叶形和叶面积应作为一个重要的表型选择指标。
研究还发现, 芽开绽期的早晚对次生代谢物含量有明显的影响; 树皮类型对京尼平甙酸、京尼平甙含量有显著影响, 而对其它4种次生代谢物含量影响不明显; 胸径生长量、树高生长量大小和硝酸还原酶活力对次生代谢物含量无显著影响; 性别对次生代谢物的合成和积累无明显影响。这些研究结果其原因待于进一步研究。
冯煦, 李鸿英. 1990. 北柴胡与烟台柴胡黄酮成分的比较研究. 中草药, 21(8): 5-6. DOI:10.3321/j.issn:0253-2670.1990.08.002 |
高锦明, 张鞍灵, 张康健, 等. 1999. 绿原酸分布、提取与生物活性研究综述. 西北林学院学报, 14(2): 73-82. DOI:10.3969/j.issn.1001-7461.1999.02.014 |
林世俊. 1985. 中药质量与道地药材———徐大椿的《药性变迁论》初探. 中药通报, 10(12): 10-11. |
马柏林, 梁淑芳, 张康健. 1996. 杜仲叶橡胶丝网络结构的研究. 西北林学院学报, 11(2): 10-13. |
马柏林, 王蓝, 张康健, 等. 1994. 杜仲胶实验室提取方法的研究. 西北林学院学报, 9(4): 67-69. |
田兰馨, 胡正海. 1983. 杜仲胶丝的形态和分布规律的研究. 西北植物研究, 3(增刊): 1-8. |
尉芹, 马希汉, 张康健. 1995. 杜仲化学成分研究. 西北林学院学报, 10(4): 88-93. |
肖小河. 1989. 中药材品质变异的生态生物学探讨. 中草药, 20(8): 42-46. DOI:10.3321/j.issn:0253-2670.1989.08.007 |
张康健, 王蓝, 马希汉. 1996. 杜仲综合开发的进展与前景. 西北林学院学报, 11(2): 75-79. |
张康健, 马希汉, 马梅, 等. 1999. 杜仲叶次生代谢物生长积累动态的研究. 林业科学, 35(2): 15-20. DOI:10.3321/j.issn:1001-7488.1999.02.003 |
张康健, 张檀. 1997. 中国神树———杜仲. 北京: 中国林业出版社, 1-17.
|
张檀, 张康健, 马惠玲, 等. 1994. 杜仲优树硝酸还原酶活力与其生长量关系的研究. 西北林学院学报, 9(4): 17-21. |
章文才主编.果树研究法(第二版).北京: 农业出版社, 1991, 456
|
Takahashi T, Matsumoto N, Oshio H. 1988. The Stability of bio-active components in the bark of Eucommia ulmoides; Eucommiae cortex. Shoyakugaku Zasshi, 42(2): 111-115. |